.png)
1.png)
2.png)
3.png)

Boletín del Instituto de Geología
Volumen 124, Núm. 1, 2022, p. 1 – 43
Guía osteológica de los dinosaurios hadrosauroideos (Ornithopoda, Hadrosauroidea), primera parte: esqueleto postcraneano
Ramírez-Velasco, Angel Alejandro1,* ; Alvarado Ortega, Jesús2
; Alvarado Ortega, Jesús2
1 Posgrado en Ciencias Biológicas, Instituto de Geología, Universidad Nacional Autónoma de México, Circuito de la investigación s/n, Ciudad Universitaria, Coyoacán, Ciudad de México, 04510.
2 Instituto de Geología, Universidad Nacional Autónoma de México, Circuito de la Investigación Científica s/n, Ciudad Universitaria, Coyoacán, Ciudad de México, 04510.
* angelalegandro@gmail.com
Resumen
Los nuevos descubrimientos de hadrosauroideos han incrementado el interés por el estudio de este grupo en México y América del Sur. En ocasiones, para los investigadores y estudiantes de habla hispana la descripción de su anatomía postcraneana suele ser una tarea complicada, debido a que el material publicado en este idioma es extremadamente escaso y los nombres de las estructuras anatómicas en inglés presentan variantes dependiendo del autor. El propósito del presente estudio es dar una guía introductoria, actualizada e ilustrada que permita reconocer los estados de carácter empleados en las filogenias de este grupo y facilite la identificación de huesos aislados o asociados, así como ofrecer una referencia de consulta en la descripción de los huesos postcraneanos para estudiantes y académicos.
Palabras clave: Anatomía, esqueleto axial, esqueleto apendicular, Hadrosauridae.
Abstract
New discoveries of hadrosauroids have increased interest in the study of this group in Mexico and South America. However, for Spanish-speaking researchers and students, describing their postcranial anatomy is often a complicated task. The material published in this language is extremely scarce and the names of anatomical structures in English have variants depending on the author. The purpose of this study is to present an introductory guide on the postcranial anatomy of hadrosauroids, updated and illustrated, which allows to recognize the states of character used in the phylogenies of this group, facilitate the identification of isolated or associated bones, as well as offer a reference guide in the description of postcranial bones for students and academics.
Keywords: Anatomy, axial skeleton, appendicular skeleton, Hadrosauridae.
Manuscrito recibido: Abril 24, 2021.
Manuscrito corregido recibido: Julio 6, 2021.
Manuscrito aceptado: Agosto 11, 2021.

1. Introducción
Los hadrosauroideos (Superfamilia Hadrosauroidea) es un grupo de dinosaurios ornitópodos que comprende a Hadrosaurus foulkii Leidy, 1858 y a todos los taxones cercanos a éste, excluyendo Iguanodon bernissartensis Boulenger, 1881 (Sereno, 1986; Figura 1). De acuerdo a Prieto-Márquez (2010), este clado está soportado por las siguientes sinapomorfías: tres dientes por alvéolo en la parte media del dentario; dentario con una inclinación ventral ubicado alrededor del 66% al 78% de la longitud de la batería dental; ausencia del foramen mandibular externo (= foramen accesorio) en el surangular; proceso premaxilar nasal extendido hasta el borde rostral de la abertura nasal ósea; rama ascendente maxilar posicionado ligeramente caudal a la mitad de su longitud; yugal con el borde orbital e infratemporal del mismo tamaño; cóndilo occipital proyectado horizontalmente; ausencia de huesos supraorbitales; y un cuerpo iliaco profundo (proporción altura/longitud mayor a 0.8).
El registro fósil de los hadrosauroideos es abundante y usualmente, bien preservado, dicho registro abarca desde el Barremiano temprano hasta el Maastrichtiano tardío en Europa, Asia, América, Antártida y África (Prieto-Márquez y Norell, 2010; Longrich et al., 2020). Anatómicamente, son de los dinosaurios mejor conocidos, debido al descubrimiento de numerosos esqueletos articulados, huesos aislados o asociados. Es por ello que su anatomía ha sido descrita e ilustrada por múltiples autores (e.g., Lambe, 1920; Lull y Wright, 1942; Weishampel y Horner, 1990), siendo el último trabajo recopilatorio el de Horner et al. (2004). Desde entonces se ha incrementado el número de especies descritas de 44 en el 2004 a 99 en el 2021 (Tabla 1), lo que plantea la necesidad de realizar nuevas recopilaciones. No obstante, para el habla hispana no existe ningún manuscrito como estos para hadrosauroideos.
El propósito de este trabajo es presentar una guía introductoria de la anatomía postcraneana de los hadrosauroideos, actualizada y acompañada de ilustraciones. Además, se espera que esta revisión sea de utilidad para aquellos estudiantes e investigadores que se enfrenten por primera vez a la descripción de hadrosauroideos en países de habla hispana.
2. Materiales y métodos
Las descripciones, comparaciones e ilustraciones de los huesos postcraneanos realizadas por el primer autor en este estudio se basan en observaciones de ejemplares depositados en las colecciones mexicanas (ver abreviaturas institucionales), así como en los descritos o ilustrados en la literatura (ver lista de referencias).
2.1. Abreviaturas institucionales
CIC, Colección Paleontológica Centro INAH (Instituto Nacional de Antropología e Historia) Coahuila, Saltillo, Coahuila; CPC, Colección Paleontológica de Coahuila, Museo del Desierto, Saltillo, Coahuila; IGM, Colección Nacional de Paleontología del Instituto de Geología, Universidad Nacional Autónoma de México, Ciudad de México; PASAC, Paleontólogos Aficionados de Sabinas, A.C., Coahuila; USNM, National Museum of Natural History, Washington, D.C (fotografías de la página web https://www.si.edu/unit/natural-history-museum).
2.2. Nomenclatura
Los nombres y términos aquí seleccionados se adoptaron de la Nomina Anatomica Avium (NAA), de acuerdo con las sugerencias de Harris (2004) para evitar términos de la anatomía mamaliana aplicada a la descripción de reptiles o aves (e.g., epipófisis vs. torus dorsal, anterior vs. craneal). Dado que algunos términos de la NAA no son usualmente aplicados por los paleontólogos (e.g., escapo, plano, torus), se creó un glosario de términos osteológicos (Tabla 2; Figuras suplementarias S1, S2 del Anexo 1) con sus respectivas definiciones obtenidas de textos especializados en anatomía ósea (Dorian, 1988; Álvarez del Villar et al., 2007; Frandson et al., 2009; Dorland, 2012; White et al., 2012). Además, debido a la gran cantidad de sinonimias y homonimias que definen determinadas estructuras anatómicas, se decidió mencionar algunas entre paréntesis en su idioma original, y crear un listado de todas ellas con sus respectivas referencias (ver Anexo 1). Para las láminas vertebrales, se sigue la terminología propuesta por Wilson (1999) y para las estructuras de la cintura pélvica se sigue a Carpenter e Ishida (2010), con las modificaciones de Ramírez-Velasco et al. (2021a).
3. Resultados
Para fines comparativos, en este trabajo, los hadrosauroideos se dividen en cuatro grupos, incluyendo dos de naturaleza parafilética: los Hadrosauroidea no-hadrosáuridos o simplemente hadrosauroideos basales, los Hadrosauridae no-euhadrosauria o hadrosáuridos basales; así como dos clados: los Lambeosaurinae o lambeosaurinos y los Saurolophinae o saurolofinos (Figura 1).
3.1. Columna vertebral
La columna vertebral de los hadrosauroideos está formada por 83 a 117 vértebras que protegen a la médula espinal (Figura 2). Se distingue en ella cuatro regiones: cervical, torácica (= dorsal), sacra y caudal. De cada región de la columna, es posible diferenciar subregiones como: anteriores, media y posteriores; se prefieren estos términos para evitar confusión al usar frases como: vértebra caudal caudal.
3.2. Vértebras
Comúnmente, las vértebras de los hadrosauroideos (Figuras 3.1–3.33), como en otros dinosaurios, están constituidas por un cuerpo vertebral cilíndrico, constreñida en su centro (llamada concavidad vertebral) y expandida hacia los extremos caudal y craneal (Figura 3.2); de esta estructura se unen de los bordes dorsolaterales un arco vertebral ligeramente laminar, que forma las paredes dorsal y laterales del foramen vertebral (= neural canal). A su vez, de este arco nace una serie de proyecciones: los procesos transversos lateralmente, un proceso espinoso dorsalmente (= neural spine) y las cigoapófisis craneales y caudales (= prezygapophysis/postzygapophysis).
En vista lateral, la base del arco vertebral (= neural arch) presenta sus bordes craneo-caudales escotados que al unirse con la vértebra contigua forman un orificio llamado foramen intervertebral, más pronunciado en la región torácica que en las otras regiones de la columna vertebral de los hadrosauroideos (Figura 3.6).
Vértebras cervicales. El número de cervicales es 11 en hadrosauroideos basales, 12–13 en saurolofinos y 13–15 en lambeosaurinos (Horner et al., 2004). Sólo Olorotitan presenta 18 cervicales (Godefroit et al., 2012a).
El atlas o primera vértebra cervical se caracteriza por poseer dos hemiarcos unidos al cuerpo del atlas (Figuras 3.7–3.9). El hemiarco atlantal en los hadrosauroideos tiene forma rectangular en vista lateral, más alta que ancha, con su borde caudal cóncavo y un borde craneal sinuoso. Cranealmente, cada hemiarco tiene una base más robusta que su parte superior laminar. De su base nace la fosa cotiloidea ovalada hacia craneal. Caudalmente, a la mitad de la altura del arco surge la cigoapófisis caudal en forma de una proyección plana, con una faceta cuadrangular ventralmente. En Altirhinus (Norman, 1998, fig. 23) presenta un proceso rostral sobre el arco, poco desarrollado o ausente en otros hadrosauroideos.
El cuerpo atlantal (= hypocentrum, intercentrum) es una estructura ovalada con el eje mayor horizontalmente en vista craneal/caudal (Figuras 3.7–3.9). Cranealmente, toda su superficie dorso-craneal forma la fosa cotiloidea y junto con las fosas del mismo nombre del arco sostienen el cóndilo occipital del cráneo.
El axis o segunda vértebra cervical es el elemento más grande de la serie cervical en los hadrosauroideos. Se caracteriza por presentar un proceso espinoso rectangular lateralmente (Figuras 3.10–3.12), cuyo borde dorsal puede ser sinuoso como en Brachylophosaurus, Maiasaura, Saurolophus e Hypacrosaurus altispinus (Figura 3.10) o generalmente convexo (Prieto-Márquez, 2008; Figura 3.11). En la cara articular craneal del cuerpo vertebral posee un proceso odontoide, de forma cilíndrica, con una superficie cóncava dorsalmente y una sección estrecha o cuello en la mitad de su longitud (Figuras 3.10–3.12). Este proceso varía en ser largo, como en la mayoría de los hadrosauroideos, o corto como en Bactrosaurus, Tanius sinensis y el hadrosauroideo de Woodbine (Main et al., 2014; Figura 3.12).
Las vértebras postaxiales tienen un cuerpo vertebral (= centrum) fuertemente opistocélico (con la cara articular craneal convexa y la cara articular caudal cóncava), con cigoapófisis craneales bajas encima del proceso transverso (= diapophysis), largos y pedunculares cigoapófisis caudales, bajos y pequeños procesos espinosos y procesos transversos cilíndricos (Horner et al., 2004; Figuras 3.13–3.19). Las cervicales anteriores suelen tener procesos transversos cortos y usualmente su cuerpo vertebral es más largo (Campione, 2014). Hacia la región posterior, el cuerpo vertebral cervical se acorta y los procesos transversos junto con los procesos espinosos se alargan (Campione, 2014; Figuras 3.18–3.19).
En los hadrosauroideos basales, las vértebras cervicales postaxiales presentan cigoapófisis caudales más cortas que las presentes en los Hadrosauridae (Figuras 3.15–3.17). Como rasgos inusuales en los hadrosauroideos, pero comunes en los saurisquios (Coria et al., 2012; McDonald e tal., 2014), Lapampasaurus presenta forámenes pneumáticos (= pleurocoels) (Figura 3.16) y Equijubus los torus dorsales (= epipophysis) sobre las cigoapófisis caudales (Figura 3.17).
Vértebras torácicas. El número de torácicas es 16 en hadrosauroideos basales y 16–20 en hadrosáuridos (Horner et al., 2004). Poseen un cuerpo vertebral más largo que las cervicales y cuentan con una cresta ventral longitudinal (Figuras 3.1–3.6, 3.20–3.33). En vista craneal, puede ser de contorno circular como en la mayoría de los hadrosauroideos basales, o bien, acorazonado como en Gilmoreosaurus, Bactrosaurus y los hadrosáuridos (Takasaki et al., 2020). A un costado y en la base del arco vertebral presenta una eminencia costolateral (= parapophysis), poco sobresaliente lateralmente y con forma ovalada (Figuras 3.4–3.6). En ella, se observa la fóvea costal (= diapophysis), una concavidad donde se une la cabeza costal de las costillas torácicas.
En el arco vertebral, nacen dos procesos transversos largos con forma de prisma triangular, fuertemente excavados hacia caudal, por la fosa infratransversal (= postzygapophyseal centrodiapophyseal fossa, transverse process fossa). Esta fosa, junto con la serie de láminas vertebrales que recorren el proceso transverso de los hadrosauroidea (Figuras 3.3–3.5), son rasgos compartidos con los dinosaurios basales, silesáuridos y pseudosuquios, sin ser considerados verdaderas evidencias de la presencia de sacos aéreos (Butler et al., 2012).
El proceso espinoso torácico es una estructura rectangular, alta y lateralmente comprimida que se proyecta dorsalmente (Figuras 2.3–2.5). Cranealmente, las torácicas medias y posteriores pueden presentar la lámina prespinal (= prezygapophysis septum), una cresta dorsoventral que nace del espacio entre las dos cigoapófisis craneales (Conti et al., 2020), acompañada o no por la fosa espinoprecigapofisial (= sulcus). Esta lámina es común observarla en los hadrosauroideos no-lambeosaurinos (Conti et al., 2020).
En los hadrosauroideos donde se conoce la columna vertebral articulada, se aprecia la transición de torácicas anteriores a posteriores (Figuras 3.21–3.22), que va de un cuerpo vertebral bajo y opistocélico, a alto y acélico (cara craneal y caudal casi planas) (Campione, 2014; Takasaki et al., 2020). Los procesos transversos varían de inclinados dorsocaudalmente a casi horizontales (Campione, 2014). Usualmente, los hadrosauroideos tienen procesos espinosos torácicos casi tan altos o incluso dos veces la altura de sus cuerpos, son raros los que sobrepasan tres veces la altura de sus cuerpos vertebrales, como ocurren en Tanius, Arenysaurus, Magnapaulia, Hypacrosaurus, Bonapartesaurus y Barsboldia (Prieto-Márquez, 2008).
En los hadrosauroideos no-lambeosaurinos, las vértebras torácicas anteriores presentan procesos espinosos anchos y rectos (Figura 3.1), mientras que en los lambeosaurinos son delgadas e inclinadas (Figura 3.6). En algunos hadrosauroideos, el extremo dorsal del proceso espinoso puede estar engrosado lateralmente, con forma espatulada cráneo/caudalmente (e.g., Tanius, Amurosaurus, Bonapartesaurus y Barsboldia) (Figuras 3.24–3.25) y raramente con una expansión de su ángulo craneal, con su mismo grosor en vista craneal (e.g., Tethyshadros y Bactrosaurus) (Figuras 3.21–3.22).
Como característica única, las vértebras torácicas de Equijubus presentan una placa central debajo de la cigoapófisis caudal, denominada hipósfeno (Figuras 3.31–3.33). McDonald et al. (2014) lo consideraron como rasgo convergente con los dinosaurios sauropodomorfos y terópodos, ya que ningún otro ornitisquio basal lo presenta.
Sacro. En los hadrosauroideos es una unidad rígida de vértebras fusionadas, compuesta de una vértebra proveniente de la serie torácica, denominada vértebra toracosacral, de cinco a siete vértebras sacras, y de la incorporación de dos vértebras caudales llamadas sacrocaudales (Figuras 4.1–4.7). El número de elementos vertebrales del sacro varía entre 6 a 8 en hadrosauroideos basales, y de 9 a 10 en Nanyangosaurus, Huehuecanauhtlus, Tethyshadros y todos los Hadrosauridae (Horner et al., 2004; Prieto-Márquez, 2008). Sólo Olorotitan posee 15 vértebras en el sacro (Godefroit et al., 2003).
En vista ventral, la vértebra toracosacral es la más ancha, de la segunda a la cuarta sacral son más angostas y se ensanchan a partir de la quinta hasta la última del sacro, siendo todas las posteriores del mismo grosor (Figuras 4.2 y 4.4). Usualmente, el sacro en vista ventral presenta un surco en su extremo caudal y cranealmente una cresta (Prieto-Márquez, 2008), no obstante, hay ocasiones donde sólo se presentan uno de los dos como es el caso de Tsintaosaurus (Figura 4.4).
En vista lateral, la toracosacral (Figura 4.5) presenta un proceso transverso cilíndrico, ya que carece de las eminencias costolaterales típicas de las vértebras torácicas. Las verdaderas vértebras sacras (Figura 4.6) poseen un proceso costal sacral (= sacral rib), conformado por una estructura cilíndrica (reminiscencia del proceso transverso) unida a una lámina sacral que se conecta con otra estructura cilíndrica ventral (reminiscencia de la verdadera costilla sacral). Sobre esta última se forma una gran placa gruesa y larga, denominada yugo sacral (= acetabular bar, yoke-like sacral bar) (Figuras 4.1–4.4). Por otro lado, las sacrocaudales (Figura 4.7) poseen procesos costales triangulares en vista craneal, en forma de placa expandida en su base y hacia su extremo lateral en forma cilíndrica. Estas últimas estructuras no se unen al ilion.
En hadrosauroideos y en los saurolofinos, la región laminar del proceso costal sacral es recta verticalmente; en cambio, en los lambeosaurinos se encuentra inclinada, craneal o caudalmente en su parte dorsal (Taylor y Lucas, 2018).
Vértebras caudales. El número de vértebras caudales en los hadrosauroideos es variable, posiblemente, los saurolofinos poseen entre 76–77, mientras los lambeosaurinos 70 (Gilmore, 1935; Godefroit et al., 2001; Dalla Vecchia, 2020). Cada región del sacro presenta sus rasgos distintivos: las anteriores se caracterizan por tener cuerpos vertebrales circulares cranealmente, procesos espinosos altos y procesos costales robustos y curvos ventralmente (Figuras 4.8–4.10); las medias presentan cuerpos vertebrales rectangulares a hexagonales cranealmente, con hemapófisis (= chevron facet, hemal arch articulation site) y procesos espinosos inclinados caudalmente (Figuras 4.11–4.16); por último, las posteriores presentan cuerpos vertebrales que van de hexagonales a circulares con o sin procesos espinales y carecen de las hemapófisis (Dalla Vecchia, 2009; Figuras 4.17–4.19).
En Tanius, Bonapartesaurus y Barsboldia, el extremo dorsal del proceso espinoso caudal es espatulado como los torácicos (Figuras 4.12–4.13). En Olorotitan, presentan una pequeña expansión craneal en su extremo dorsal (Figura 4.11). En Tethyshadros, tienen una forma espatulada (Figura 4.19). Finalmente, en Arenysaurus, presentan una base esbelta que se engrosa progresivamente hacia su extremo dorsal (Figura 3.20). La mayoría de los lambeosaurinos poseen la fosa espinoprecigapofisial (= sulcus) en la base del proceso espinoso, ésta se extiende hasta la mitad de la altura del proceso, rasgo inusual en los hadrosauroideos no-lambeosaurinos (Conti et al., 2020; Figura 4.21).
3.3. Costillas
Son huesos planos y alargados, situados en las partes laterales de las vértebras cervicales y torácicas (Horner et al., 2004). Todas las costillas tienen un cuerpo y dos extremos (Figura 5.1). Su cuerpo costal es aplanado transversalmente, con una cara medial cóncava y una lateral convexa. El extremo dorsal está bifurcado, al presentar la cabeza (= capitulum) unida al cuello y la tuberosidad costal (= tuberculum). Por último, el extremo ventral costal puede ser una terminación redondeada o rugosa, la cual recibe el cartílago costal (Figuras 5.9 y 5.10).
Costillas cervicales. Son las más cortas de todas (Figuras 3.13, 3.14, 5.1). Su cuello y cabeza costal están deprimidos y orientados hacia medial; en cambio, la tuberosidad costal, larga y cilíndrica, se orienta hacia dorsal. Su extremo ventral costal es corto, acuminado y aplanado dorsoventralmente.
Costillas torácicas. Son alargadas, con un ángulo costal amplio (Figura 5.2), siendo las más largas las que están cercanas a la escápula (Campione, 2014). Su extremidad dorsal está comprimida craneocaudalmente y presentan un cuello costal más largo que las cervicales. En cambio, progresivamente, su cuerpo costal se va aplanando lateromedialmente y provoca el desarrollo de una cresta costal hacia caudal a todo lo largo de su longitud. En las costillas torácicas más caudales se pierde esta cresta (Campione, 2014).
3.4. Cheurón
En hadrosauroideos son huesos alargados, planos lateralmente y con forma de “Y” (Figuras 5.4–5.8). Se componen de dos secciones: el arco hemal o la parte superior bifurcada, con la que se une a los cuerpos vertebrales caudales; y la espina hemal, la parte inferior elongada y quillada caudoventralmente. Usualmente, la espina hemal es recta respecto al arco hemal lateralmente. Sólo en Tethyshadros, la espina hemal presenta una expansión en forma de bota distalmente (Figuras 5.7–5.8), rasgo compartido con los dinosaurios terópodos tetanuros (Dalla Vecchia, 2020).
3.5. Tendones osificados
Son estructuras largas y aciculares que surgen de la osificación natural de los tendones epiaxiales (Organ y Adams , 2005). De acuerdo con Prieto-Márquez (2007), en los hadrosauroideos, estos son de cilíndricos a comprimidos como una cinta (Figuras 5.9, 5.10). Se localizan a los costados de los procesos espinosos de la región torácica, sacra y caudal de la columna vertebral de los hadrosauroideos (Organ y Adams, 2005). En Brachylophosaurus posee tendones en los costados del extremo dorsal de las costillas torácicas, lateroventralmente de los procesos transversos torácicos y en la base de los procesos costales caudales (Prieto-Márquez, 2007).
Los tendones osificados en los hadrosauroideos se extienden desde la cuarta vértebra torácica hasta la sexta caudal (e.g., Bolong y Brachylophosaurus) o pueden alcanzar la vértebra caudal 20 (e.g., Tethyshadros, Corythosaurus y Gryposaurus) (Wu y Godefroit, 2012). A diferencia de los iguanodóntidos, los hadrosauroideos carecen de tendones osificados en la región cervical (Wu y Godefroit, 2012).
Entre los hadrosauroideos, los tendones osificados están dispuestos de dos maneras, paralela o formando un arreglo reticular (Dalla Vecchia, 2009; Wang et al., 2011; Wu y Godefroit, 2012). Por ejemplo, en Bolong, presenta un patrón paralelo en todo su cuerpo; en Jinzhousaurus, es paralelo predominante pero reticulado entre la torácica 10–11; en Tethyshadros, nuevamente es paralelo predominante con un reticulado entre las caudales 1–6. Por último, los tendones osificados de los hadrosáuridos son enteramente reticulados.
3.6. Cintura pectoral
Esta región se forma por tres huesos pares articulados entre sí. Estos son: el coracoides, la escápula y la placa esternal. Tanto la escápula como el coracoides conforman la cavidad glenoidea, al unir ambas caras articulares humerales de cada hueso (Figura 6.1).
Coracoides. En los hadrosauroideos es el elemento más pequeño de la cintura pectoral (Figuras 6.1–6.7). Lateralmente, es un hueso con forma de gancho comprimido hacia craneoventral, con dos superficies articulares rectas dorsocaudal y ventrocaudalmente. Se compone de dos secciones, el cuerpo coracoideo y el extremo esternal.
El cuerpo coracoideo ocupa la parte caudal del hueso, tiene una cara lateral cóncava y la medial convexa (Figuras 6.2–6.3). Dorsalmente, la parte caudal es más gruesa que la craneal. En vista caudolateral, presenta una superficie dorsocaudal plana y rugosa, ahí se une con la escápula. Debajo de ella, hay una superficie rugosa cóncava, donde se articula con el húmero (= glenoid). En la región central o cerca del borde caudal, está perforada de forma circular por el foramen del nervio supracoracoideo (= coracoid foramen). En el borde dorsal del cuerpo, sobresale lateralmente el tubérculo del bíceps (= lateral process, bicipital knob), una estructura semiesférica redondeada donde se inserta el músculo bíceps braquial (Dilkes, 2000).
El extremo esternal (= hook-like process, ventral hook, ventral process, sternal blade) es la estructura en forma de gancho del coracoide, proyectado ventralmente (Figuras 6.2–6.7). Dorsalmente, presenta una cresta baja o cresta articular esternal que nace desde su punta ventral hasta la parte craneal del tubérculo del bíceps. En vida, la punta del extremo esternal se articula con la placa esternal correspondiente.
En los hadrosauroideos más basales, el cuerpo coracoideo tiene un borde dorsal convexo o ligeramente cóncavo (Figuras 5.4–5.5), un tubérculo poco desarrollado, una cara articular escapular más alta que la fosa glenoidea y un extremo esternal pequeño (Prieto-Márquez, 2008; Figuras 6.4–6.5). En cambio, en los Euhadrosauria, el cuerpo coracoideo tiene un borde dorsal cóncavo, un tubérculo del bíceps prominente, una fosa glenoidea más alta que la cara articular humeral y un extremo esternal largo (Brett-Surman y Wagner, 2007; Prieto-Márquez, 2008) (Figuras 6.2–6.3, 6.6–6.7). Este extremo, dentro de los hadrosáuridos varía desde una forma robusta, casi tan largo como alto (e.g., Edmontosaurini, Kritosaurini, Parasaurolophini y la mayoría de los Lambeosaurini) (Figuras 6.2, 6.4–6.5) a una forma angosta, más alto que largo (e.g., Arenysaurus, Amurosaurus, Wulagasaurus, Brachylophosaurus y Saurolophini) (Figuras 6.6–6.7). En Latirhinus el coracoide es una combinación de ambos morfotipos, un cuerpo de los hadrosauroideo basales con el extremo esternal largo típico de los euhadrosaurios.
Escápula. Es un hueso largo, aplanado lateromedialmente y ligeramente curvado hacia medial y ventralmente (Figuras 6.1, 6.8–6.16). Se reconocen tres regiones: el extremo craneal escapular, el cuello y el cuerpo escapular (Brett-Surman, 1989; Brett-Surman y Wagner, 2007).
El extremo craneal (= proximal part) es la región más amplia y ancha de la escápula, puesto que presenta la superficie articular coracoidea cranealmente, la superficie articular humeral craneoventralmente y tres procesos (Figura 6.8). La cara medial de esta región es plana y la lateral cóncava, debido a la fosa deltoidea (Figuras 6.8–6.10). El proceso glenoideo (= ventral protuberance, scapular labrum, glenoid apex) es una extensión triangular ventral de esta región, de donde se inserta el músculo tríceps braquial y de la cual forma parte de la superficie articular humeral caudal (Dilkes, 2000). Del borde dorsolateral de la región, nacen dos estructuras, el acromion (= pseudoacromion process), una proyección triangular en vista dorsal, y la cresta deltoidea, una cresta baja que se origina del borde caudal del acromion (Prieto-Márquez, 2008).
El cuello escapular (= neck, proximal constriction) es la región que se constriñe en vista lateral, entre el extremo y el cuerpo escapulares (Figura 6.9). Puede ubicarse caudal a la cresta deltoidea o a la mitad de ésta.
El cuerpo escapular (= blade, scapular blade) es la parte rectangular más larga de la escápula (Brett-Surman y Wagner, 2007; Figuras 6.9, 6.10). Usualmente, esta región equivale tres veces la altura del extremo craneal escapular. En Bolong, Altirhinus, Tethyshadros y Adynomosaurus (Figura 6.16), la región es significantemente más corta, pues representa dos veces la altura del extremo craneal (Prieto-Márquez et al., 2019b). El extremo caudal del cuerpo está expandido dorsoventralmente, aunque entre los Lambeosaurinae, Brachylophosaurini, Saurolophini y Edmontosaurus, dicha expansión es más conspicua que en el resto del clado (Prieto-Márquez, 2008).
Dentro de los hadrosauroideos, se pueden reconocer tres morfologías (Prieto-Márquez, 2008): escápulas rectas con un proceso acromion curvado dorsalmente (e.g., hadrosauroideos basales) (Figuras 6.1, 6.11); escápulas con su extremo craneal inclinada ventralmente y con un acromion recto (e.g., saurolofinos) (Figuras 6.9, 6.14); y escápulas igualmente inclinadas ventralmente, pero con un acromion curvado dorsalmente (e.g., lambeosaurinos) (Figuras 6.12–6.15).
Placa esternal. Es un hueso con forma de hacha (Figuras 6.17–6.23). Se compone del cuerpo esternal y del proceso caudolateral (Brett-Surman, 1989).
El cuerpo esternal (= sternal plate, sternal blade, medial flange, craniomedial flange) es la región más ancha y tiene forma triangular en vista dorsal/ventral (Figura 6.20). El borde medial es fuertemente convexo y rugoso, el borde lateral va de recto a ligeramente cóncavo y su borde caudal es cóncavo. Puede presentar o no un proceso caudomedial con forma de gancho. Las placas se unen a través de sus bordes mediales y se articulan con el coracoides desde el ángulo craneal o ápice.
El proceso caudolateral (= sternal handle) es una estructura alargada, cilíndrica, ligeramente aplanada (Figura 6.19). Su extremo caudal tiene una superficie rugosa, de donde se unirían costillas cartilaginosas.
Entre los hadrosauroideos se conocen dos morfologías (Prieto-Márquez, 2008): placas esternales con un proceso caudolateral de la misma longitud que su cuerpo esternal (e.g., hadrosauroideos basales, Adynomosaurus, Amurosaurus y Tsintaosaurus) (Figuras 6.21–6.22); o placas esternales con un proceso caudolateral más largo que su cuerpo (e.g., hadrosáuridos, Tethyshadros, Gobihadros y Probactrosaurus) (Figuras 6.17, 6.20, 6.23).
3.7. Miembro torácico
Esta región corporal se compone del húmero o hueso del brazo, del radio y la ulna o huesos del antebrazo, así como de huesos de la mano que incluyen el carpo, el metacarpo y las falanges manuales.
Húmero. Es largo con una pronunciada cresta triangular; se distingue en él un cuerpo y dos extremos (Figuras 7.1–7.4).
El cuerpo humeral (= humeral shaft) es el eje del hueso, tiene forma cilíndrica y ahí nacen todos los procesos. El extremo proximal es expandido y el distal es angosto, denominado cuello humeral (Figura 7.1). En vista caudal, está una superficie rugosa alargada o línea del músculo del dorsal ancho dentro de la base del extremo proximal (Figuras 7.1–7.2, 7.4).
El extremo proximal del húmero muestra la cabeza humeral como una superficie articular semiesférica, proyectada principalmente en la superficie dorsocaudal y con una pequeña extensión en la superficie craneal (Figuras 7.3, 7.4). Cranealmente, en la cabeza humeral se observa el tubérculo lateral (= outer tuberosity, tuberculum dorsale), una superficie rugosa redondeada de donde nace la cresta deltopectoral (= radial crest, deltoid crest) distalmente (Figuras 7.1–7.3). Ésta es triangular verticalmente en vista lateral, con su extremo más angosto hacia proximal y la más amplia distalmente. Tiene una superficie lateral convexa y una medial cóncava, con un ápice rugoso distalmente inclinado medialmente (Figuras 7.1, 7.3, 7.4). En el lado opuesto a esta cresta, se ubica el tubérculo medial (= inner tuberosity, tuberculum ventrale), una superficie rugosa redondeada donde nace la cresta del bíceps distalmente, de menor longitud que la cresta deltopectoral (Figuras 7.1, 7.3–7.4). Cranealmente, entre las dos tuberosidades, está el plano intertuberal, un área plana y lisa delimitada por la cresta deltopectoral lateralmente y la cresta del bíceps caudalmente (Figuras 7.1, 7.3).
El extremo distal humeral se distingue por su terminación bicondilar (Figuras 7.2–7,4, 7.6). Las caras articulares de los cóndilos son rugosas; en cambio, las externas e internas son lisas y rectas. La medial del cóndilo medial presenta una superficie cóncava denominada epicóndilo medial (= epicondylus dorsalis, entepicondylar groove) (Figura 7.1). En vistas craneal y caudal, los cóndilos están separados por dos fosas triangulares que convergen en la vista distal. Hacia craneal, la fosa braquial es relativamente más pequeña, y en vista caudal, la fosa para el olécranon (= caudal groove) es de mayor tamaño (Figuras 7.3, 7.4, 7.6).
En los hadrosauroideos hay tres tipos de húmeros (Brett-Surman y Wagner, 2007; Prieto-Márquez, 2008, 2011). En los hadrosauroideos basales y hadrosáuridos basales, el húmero cuenta con una cresta deltopectoral corta y angosta, menor que el 48% de la longitud total del hueso (Figuras 7.7, 7.11). En los saurolofinos existe una cresta deltopectoral larga, entre el 48 al 55%, ligeramente expandida cranealmente (Figuras 7.1–7.2, 7.9). Por último, en los lambeosaurinos, el húmero tiene una cresta deltopectoral amplia y larga, superior al 55% de su longitud total, lo que les da una apariencia más robusta (Figuras 7.8, 7.10).
Ulna. Es un elemento largo con una terminación proximal expandida (Horner et al., 2004). Como todo hueso largo, se compone de un cuerpo y dos extremos (Figuras 7.12–7.19).
El cuerpo de la ulna en los hadrosauroideos tiene una forma de prisma triangular (Figuras 7.12–7.16). Las caras lateral y medial son ligeramente convexas, la craneal, lisa y redondeada. Únicamente Saurolophus presenta un cuerpo ulnar cilíndrico (Horner et al., 2004). De sus bordes, el caudal es el más conspicuo porque forma una cresta longitudinal delgada, la cual se conecta proximalmente con el olécranon, y se extiende distalmente cerca del extremo distal, desviándose medialmente (Figura 7.12).
El extremo proximal ulnar presenta una proyección cónica característica, denominada olécranon (Figuras 7.12–7.15). La superficie craneal se articula con el húmero, área denominada escotadura del olécranon (= sigmoidal gutter, proximal radial facet) (Brett-Surman y Wagner, 2007). Distal a este proceso, se proyectan dos crestas, una lateral y otra medial (= lateral flange/medial flange), que encierran una superficie articular cóncava, triangular en vista craneal, denominada escotadura radial (= radial surface) (Brett-Surman y Wagner, 2007). Ambas crestas son de forma triangular, siendo más larga y prominente la cresta medial (Figura 7.14). La cresta lateral se distingue por presentar una superficie convexa; en cambio, la cresta medial tiene una depresión triangular o impresión braquial (Figuras 7.12, 7.14).
El extremo distal ulnar es más aplanado lateromedialmente con respecto a su cuerpo, con una superficie distal redondeada y una superficie medial rugosa o depresión radial (= distal radial facet) (Figuras 7.12–7.14, 7.16).
En la mayoría de los hadrosauroideos, la ulna es delgada y alargada, mientras que en los hadrosauroideos basales como Jinzhousaurus, Bolong, Altirhinus y algunas formas derivadas (e.g., Parasaurolophus, Bonapartesaurus, Gryposaurus latidens y Saurolophus) es comparativamente más robusta y corta. Además, las ulnas de algunos hadrosauroideos basales se distinguen por tener un extremo distal más expandido dorsoventralmente, tan alto como su extremo proximal (Figuras 7.17–7.18). En contraste, en los hadrosáuridos, el extremo distal es más estrecho que su extremo proximal (Figuras 7.12, 7.19).
Respecto a su longitud relativa con el húmero, se presentan tres proporciones (Prieto-Márquez, 2008; Brownstein y Bissell, 2020); en los hadrosauroideos no euhadrosauria, la ulna es más corta o del mismo tamaño que el húmero; en los saurolofinos y Parasaurolophus, es ligeramente más largas que el húmero; en cambio, en los lambeosaurinos y Brachylophosaurus, es mucho más larga que su húmero.
En cuanto a la forma del borde caudal (en vista lateral), los hadrosauroideos basales y la mayoría de los lambeosaurinos presentan un borde recto y elongado (Figuras 7.17, 7.19). En cambio, los saurolofinos, Parasaurolophus y Amurosaurus presentan un borde sinuoso y corto (Figuras 7.12, 7.14), demarcado por la expansión craneoventral de la región proximal (Brownstein y Bissell, 2020).
Radio. Es largo, delgado, recto y ligeramente más corto que la ulna (Lambe, 1920; Horner et al., 2004). Presenta un cuerpo y dos extremos (Figuras 7.20–7.27).
El cuerpo radial es una estructura larga casi cilíndrica (Figuras 7.20–7.21). El extremo proximal radial, denominado cabeza radial, tiene forma de cono invertido, con la parte constreñida dirigida distalmente (Figuras 7.20–7.22). La cara craneal es ligeramente convexa, mientras que la caudal es plana para la articulación con la ulna. La superficie proximal de la cabeza presenta la cótila humeral (Figura 7.22), faceta articular con la que se une al cóndilo lateral del húmero (Senter, 2012).
El extremo distal radial, más voluminoso que el proximal, tiene forma triangular en vista lateral/medial (Figuras 7.20–7.21, 7.23). El borde craneomedial es convexo y el caudolateral es de recto a sinuoso, debido a la presencia de una superficie medial de ligeramente cóncava a plana. La cara distal es una faceta articular redondeada, denominada cara articular radiocarpal (Figuras 7.20–7.21, 7.23).
En los hadrosauroideos basales, ambos extremos radiales son más grandes y robustos que en sus parientes más derivados, principalmente, es en el extremo distal donde presentan una expansión dorsoventral notable (Figura 7.25). Un rasgo distintivo de Gobihadros y los hadrosáuridos es la presencia de una línea intermuscular (= proximal keel, caudal keel, ventral ridge, interosseous ridge) que recorre toda la longitud del hueso (Figuras 7.20, 7.26). Dicha línea tiene forma de quilla cerca del extremo proximal, se extiende desde el borde medial del extremo proximal, se arquea lateralmente en el cuerpo y termina en el borde medial del extremo distal (Godefroit et al., 2012c; Conti et al., 2020; Tsogtbaatar et al., 2019).
Carpo. En los hadrosauroideos, está compuesto de 2 a 7 carpianos pequeños con formas que varían entre la discoidal, tetraédrica, oval y prismática rectangular (Figuras 8.1–8.3).
Se reconocen tres morfologías del carpo. El primer morfotipo está representado por Jinzhousaurus (Figuras 8.1, 8.5), donde consta de tres elementos: el de mayor tamaño es el carpiano fusionado radial-central, con forma de prisma rectangular; y dos carpianos pequeños discoidales, el ulnar y el distal (Wang et al., 2011). Esta morfología del carpo recuerda a la de los Iguanodontidae (Wang et al., 2011).
El segundo morfotipo está en Bolong y Altirhinus (Figura 8.2). Se integra por seis elementos casi del mismo tamaño, los cuales nunca se fusionan durante su ontogenia, como lo demuestra el individuo juvenil y adulto de Bolong (Wu y Godefroit, 2012; Zheng et al., 2014). El carpiano radial es elongado mediolateralmente con una superficie convexa distal (Wu y Godefroit, 2012). El carpiano ulnar es robusto y redondeado en sus caras (Norman, 1998, fig. 29.D). El carpiano central (= intermedium carpal) y los distales 2–5 son aplanados con una cara convexa y otra cóncava (Norman, 1998; Wu y Godefroit, 2012). Esta morfología es similar a la de los ornitópodos basales.
La tercera morfología está reconocida en los hadrosáuridos, Tethyshadros y posiblemente Gobihadros (Figuras 8.3–8.4, 8.8). Está compuesta por dos elementos (Brett-Surman y Wagner, 2007; Prieto-Márquez, 2007), aunque usualmente sólo se preserva el de mayor tamaño. El carpiano pequeño es de forma esférica a ovoide; en cambio, el carpiano de mayor tamaño tiene forma de tetraedro.
Metacarpo. En los hadrosauroideos, el metacarpo está formado por 4 o 5 metacarpianos (Figuras 8.2, 8.4–8.8). Como todo hueso largo, tienen un cuerpo y dos extremos, excepto el primer metacarpiano (Figuras 8.9–8.16).
El primer metacarpiano, cuando está presente, es un disco plano incorporado al carpiano radial, ligeramente separados por un pequeño surco (Figuras 8.1, 8.2, 8.5). Este hueso sólo es conocido en Jinzhousaurus y Bolong (Wang et al., 2011; Wu y Godefroit, 2012).
El segundo metacarpiano tiene un cuerpo semicilíndrico recto y comprimido transversalmente (Figuras 8.9–8.10). La cara lateral es plana y estriada, con su cara medial convexa. El extremo proximal está más expandido transversalmente que la distal. En los hadrosauroideos basales, el segundo metacarpiano presenta una superficie articular distal convexa, llamada tróclea (= caudodistal furrow), a diferencia de los hadrosáuridos que muestran una superficie casi plana. En Bolong y Jinzhousaurus, su extremo distal está expandido transversalmente (Figuras 8.2, 8.5) y en Eolambia, su cuerpo está ligeramente curvado medialmente (Figura 8.9).
El tercer metacarpiano tiene un cuerpo recto, largo y con forma de prisma rectangular (Figuras 8.11–8.12). El extremo distal es angosto y alto, mientras que su extremo proximal es deprimido y expandido transversalmente. En los hadrosauroideos basales, este metacarpiano presenta una tróclea digital, en lugar de la cara articular convexa de los hadrosáuridos (McDonald et al., 2012). Sólo Jinzhousaurus y Bolong presentan un metacarpiano corto y robusto, similar al de los iguanodóntidos (Figuras 8.2, 8.5).
El cuarto metacarpiano tiene un cuerpo curvado lateralmente y un extremo proximal fuertemente expandido transversalmente (Prieto-Márquez, 2001; McDonald et al., 2012; Figura 8.13). Este extremo, con forma triangular proximalmente, presenta una cara dorsal y palmar cóncavas y una medial plana. En Bolong y Jinzhousaurus, el extremo proximal es recto y su extremo distal está ligeramente expandido transversalmente, además de presentar una tróclea digital (Wang et al., 2011; Wu y Godefroit, 2012).
En los hadrosauroideos, el quinto metacarpiano puede presentar dos formas (Prieto-Márquez, 2008; Figuras 8.14–8.16). En los hadrosauroideos basales es largo, cilíndrico, recto, con extremos proximal y distal ligeramente expandidos transversalmente (McDonald et al., 2012; Figura 8.14). En los hadrosáuridos tiene un aspecto de pirámide, con su extremo distal estrecho y ligeramente torsionado dorsolateralmente (Prieto-Márquez, 2008; Figuras 8.15–8.16). Con respecto a la longitud de este metacarpiano, es variable dentro del grupo. En Bolong, Jinzhousaurus, Plesiohadros y Parasaurolophini es corto, mientras que en Eolambia, Gobihadros y la mayoría de los Hadrosauridae es largo (Prieto-Márquez, 2008).
Falanges manuales. Los huesos de los dedos se pueden subdividir, dependiendo de su posición, en falanges proximales (II-1, III-1, IV-1 y V-1), medias (II-2, III-2, IV-2 y V-2) y distales (I-1, II-3, III-3, IV-3 y V-3). La fórmula falangeal varía de 2/3/3/3/2 en Bolong, 1/3/3/2/2 en Bactrosaurus, y 0/3/3/3/3 en Tethyshadros y los Hadrosauridae (Horner et al., 2004; Dalla Vecchia, 2009). En algunos hadrosáuridos, la fórmula puede cambiar de 0/3/3/2/3 (e.g., Edmontosaurus) a 0/3/3/3/4 (e.g., Brachylophosaurus) (Prieto-Márquez, 2014; Prieto-Márquez, 2007). Al ser huesos largos, están provistos de un cuerpo y dos extremos (Figuras 8.17–8.25).
Si el dígito I está presente, la falange-1 es un disco oval (Figura 8.5), mientras que la falange-2 es una estructura cónica, recta y ligeramente comprimida mediolateralmente, semejante a la de los iguanodóntidos (Figura 8.5–8.6, 8.22). La presencia de este dígito está confirmada en los hadrosauroideos basales Bolong (Figura 8.2), Jinzhousaurus (Figura 8.5), Altirhinus, Batyrosaurus, Gobihadros, Probactrosaurus (Figura 8.6) y Eolambia (Figura 8.22) (Norman, 1998, 2002; Wang et al., 2011; McDonald et al., 2012; Godefroit et al., 2012c; Wu y Godefroit, 2012; Tsogtbaatar et al., 2019).
Las falanges manuales no-ungueales de los dígitos II-V presentan cuatro morfotipos reconocibles en vista dorsal (Figuras 8.17–8.21): cuadrangulares (Figura 8.17), rectangulares (Figuras 8.18–8.19), cilíndricas (únicas del dígito V; Figura 8.19) y cuneiformes (Figura 8.21). Cada una de ellas se distribuye de forma distinta, según el nivel taxonómico. Por ejemplo, la falange III-1 (dígito III, número uno) con forma cuadrangular es común en los hadrosauroideos basales y los lambeosaurinos (Figuras 8.2, 8.5–8.7), mientras que, en los saurolofinos, son rectangulares (Figura 8.4, 8.8). Por otro lado, la presencia de falanges cuneiformes en el dígito II y III es característico de una mano de hadrosáurido (Figuras 8.4, 8.7, 8.8).
Las ungueales de los dígitos II y III son estructuras deprimidas (Figuras 8.23–8.25). La ungueal II es asimétrica y suele ser más larga que la ungueal III (Figura 8.24). Por el contrario, la ungueal III es simétrica y más ancha que la II (Figura 8.25). En los hadrosauroideos basales, tienden a ser más largas, más agudas y con un surco lateral (Figura 8.23). En cambio, en los hadrosáuridos, tienen una forma de medialuna en vista dorsopalmar y carecen del surco lateral (Prieto-Márquez, 2008; Figuras 8.24–8.25).
3.8. Cintura pélvica
Está constituida por tres elementos: el ilion, el pubis y el isquion (Figura 9.1). La conjunción de estos huesos forma el acetábulo, una concavidad profunda con una pared interna perforada, denominada foramen acetabular (Figura 9.1), rasgo común de los dinosaurios.
En los hadrosauroidea, se abren dos agujeros por debajo del acetábulo, denominados foramen obturador y fenestra isquiopúbica (Figura 9.1). El foramen obturador, ancestralmente presente en los Sauropsida, se localiza craneoventralmente en el pubis, como un agujero ovalado o una escotadura ovalada (Hutchinson, 2001). La fenestra isquiopúbica (erróneamente identificada como foramen obturador) es un espacio alargado formado por el borde dorsal del pubis y el borde ventrocraneal del isquion, algunas veces dividido por el proceso obturador (Figura 9.1). De acuerdo con Brett-Surman y Wagner (2007), la fenestra isquiopúbica puede estar completamente osificada en algunos individuos de hadrosáuridos y dejar libre un pequeño foramen central, aquí nombrado foramen isquiático.
Ilion. Es plano lateromedialmente, alargado, compuesto por un centro o cuerpo iliaco de donde nacen dos proyecciones alares, una caudal y otra craneal (Figuras 9.2–9.19).
El ala preacetabular (= anterior process, preacetabular process, cranial process) es una proyección alargada craneal, de forma acicular, comprimida, con los bordes dorsal y ventral afilados (cresta iliaca dorsal y ventral; Figuras 9.2–9.5). Algunos hadrosauroideos basales presentan una cresta deltada o cresta preacetabular (= horizontal boot) en su borde ventral, cerca del extremo craneal del ala (e.g., Altirhinus, Xuwulong, Eolambia, Gobihadros y Huehuecanauhtlus) (McDonald et al., 2012; Ramírez-Velasco et al., 2012; Tsogtbaatar et al., 2019; Figura 9.7). Sobre su cara medial se observa el área articular vertebral (= medial sacral ridge, medial shelf), una larga cresta robusta que se extiende caudalmente desde la mitad de la longitud del ala, hasta el cuerpo iliaco. En ella se unen los procesos costales del sacro. Esta estructura puede ser gruesa si nace del borde dorsal del ala (e.g., hadrosauroideos basales) (Figura 9.10) o como una cresta delgada cuando surge a la mitad de la altura del ala (Prieto-Márquez, 2008; Figura 9.4).
El cuerpo iliaco (= iliac plate, main plate, central plate) es una placa rectangular, alta, ancha, con un borde dorsal de recto a convexo y con un borde ventral cóncavo (Figuras 9.2–9.5). La cara medial es cóncava en su mitad caudal y convexa cranealmente. La cara lateral es ligeramente cóncava (fosa iliaca dorsal) e interrumpida por dos proyecciones laterales en su región caudal: el proceso lateral iliaco y el antitrocánter (Figura 9.2). Del borde dorsal sobresale el proceso lateral iliaco (= antitrochanter, suprailiac crest, iliac boss, supratrochanteric crest, supraacetabular process, laterally everted rim), proyección con forma variada, desde sinuosa (Figuras 9.6–9.8), triangular (Figura 9.9), acampanado (Figuras 9.1–9.2, 9.12–9.13, 9.16–9.17), linguliforme (9.11, 9.14, 9.18–9.19) o trapezoidal (Figura 9.15).
El borde ventral del cuerpo se compone de dos pedúnculos en sus extremos y de la escotadura acetabular entre ellos. El pedúnculo púbico (= pubic process), ubicado cranealmente, es de forma casi cónica, delimitado ventralmente por una cresta longitudinal o cresta supracetabular (= supra-acetabular flange; Figura 9.2). Caudalmente a la escotadura acetabular, existe un abultamiento rectangular, denominado pedúnculo isquiático (= ischial process; Figuras 9.2, 9.4–9.5). En la mayoría de los hadrosauroideos, excepto Jinzhousaurus, Equijubus y Probactrosaurus (Prieto-Márquez, 2008), este pedúnculo desarrolla una proyección lateral ovalada, con una cresta central, denominada en este trabajo antitrocánter (= ischial tuberosity, caudal tuberosity, postacetabular ridge, posteroventral protuberance; Figuras 9.2, 9.4–9.5).
El ala postacetabular (= posterior process, postacetabular process) es una proyección caudal plana, triangular (Figuras 9.6–9.8, 9.11) u oblonga (Figuras 9.2, 9.9, 9.12–9.19). En vista dorsal, el ala varía de ser delgada a tan ancha como la región caudal del cuerpo iliaco (Prieto-Márquez, 2008; Figuras 9.2–9.5). La cara lateral es ligeramente cóncava (concavidad infracristal), mientras que la cara medial varía de ser plana a cóncava. En vista lateral, la superficie dorsal y lateral están delimitadas por una cresta que se origina del extremo caudal del proceso lateral iliaco, llamado cresta dorsolateral iliaco (Figuras 9.2–9.3). Su borde ventral presenta una depresión larga en forma de canal, denominado fosa breve (superficie donde se inserta el músculo caudofemoral breve), el canal puede ser visible parcialmente (e.g., Gilmoreosaurus, Equijubus y Probactrosaurus) (Figura 9.9–9.10) o totalmente en vista lateral (e.g., Jinzhousaurus, Xuwulong y Zuoyunlong) (Prieto-Márquez, 2010; You et al., 2011; Wang et al., 2015; Figura 9.6) y parcial o totalmente en vista medial (e.g., Huehuecanauhtlus, Eolambia, Tanius, Lophorhothon, Claosaurus y los miembros de la familia Hadrosauridae) (Prieto-Márquez y Salinas, 2010; Ramírez-Velasco et al., 2012; Cruzado-Caballero y Powel, 2017; Figuras 9.4, 9.10).
Se reconocen siete morfologías del ilion en todo el clado Hadrosauroidea, las cuales comenzaron a ser reconocidas por Brett-Surman y Wagner (2007) (Tabla 3). El primer morfotipo está representado por Jinzhousaurus, Eolambia y Equijubus (Figuras 9.6–9.7). Se distingue por tener un cuerpo con el borde dorsal de recto a ligeramente convexo, con un ala preacetabular de forma triangular, un cuerpo iliaco más alto que largo, así como un proceso lateral alargado y sinuoso.
El segundo morfotipo (= morfotipo uno de Brett-Surman y Wagner, 2007) está ilustrado por Gilmoreosaurus, Tanius y Claosaurus (Figuras 9.8–9.10). Es un ilion más largo que alto, con el ala postacetabular variable entre triangular a oblonga, un proceso lateral sinuoso a cuneiforme y un antitrocánter incipiente.
El tercer morfotipo (= morfotipo dos de Brett-Surman y Wagner, 2007), ejemplificado por Tethyshadros, Lophorhothon, Edmontosaurini y Saurolophus angustirostris (Figuras 9.11–9.12), es comparativamente más bajo y largo que en los anteriores, presenta un ala postacetabular oblonga y un proceso lateral entre acampanado y linguliforme.
El cuarto morfotipo (= morfotipo tres de Brett-Surman y Wagner, 2007) está representado en Hadrosaurus y los miembros de las tribus Kritosaurini y Saurolophini (Figuras 9.13–9.14). Su cuerpo es tan alto como largo, su ala postacetabular es oblonga y ligeramente inclinada dorsalmente, mientras que su proceso lateral puede ser entre acampanado y linguliforme.
El quinto morfotipo es observado sólo en los Brachylophosaurini (Figura 9.15), en los cuales el cuerpo del ilion es más alto que largo, mientras que su proceso lateral es trapezoidal o casi rectangular.
El sexto morfotipo (= morfotipo cuarto de Brett-Surman y Wagner, 2007), se encuentra representado por los Lambeosaurini y Charonosaurus (Figuras 9.1–9.2, 9.16). El ilion es más alto que largo, pero comparativamente más elongado que en el cuarto y quinto morfotipos. Su proceso lateral es acampanado y usualmente, más pequeño que en los anteriores morfotipos.
El séptimo morfotipo (= morfotipo quinto de Brett-Surman y Wagner, 2007) es ejemplificado por Parasaurolophus, Hypacrosaurus altispinus y Tsintaosaurus (Figuras 9.17–9.19). Es robusto, con el cuerpo más alto que largo, con las alas pre y postacetabular peculiarmente profundas y el proceso lateral linguliforme y alto.
Pubis. En los hadrosauroideos es plano, de forma espatulada o de gota, con la región craneal expandida dorsoventralmente (Figuras 10.1–10.11). Se compone de un cuerpo y dos proyecciones en cada extremo, el escapo púbico y el ala prepúbica (Baumel y Witmer, 1993; Figuras 10.1–10.3).
El cuerpo púbico es la región caudal del hueso, tiene forma rectangular vertical y de borde caudal cóncavo, denominado escotadura acetabular púbica (Figuras 10.1–10.2). Su cara lateral se distingue por presentar una cresta acetabular (= acetabular margin) paralela a la escotadura, la cresta puede ser tan alta como ésta (e.g., la mayoría de los hadrosauroideos basales) (Figura 10.4) o sólo estar presente en la mitad dorsal del acetábulo (e.g., Hueheucanauhtlus, Eolambia, Lophorhothon, Eotrachodon y los Hadrosauridae) (Figuras 10.1, 10.5–10.11). Del cuerpo púbico nacen dos pedúnculos, el iliaco dorsalmente y el isquiático caudoventralmente. El pedúnculo iliaco (= dorsal process, iliac process) es una proyección trapezoidal plana, mientras que el pedúnculo isquiático (= ischial process) oscila entre una estructura plana cuadrangular (e.g., hadrosauroideos basales) y cilíndrica (e.g., hadrosáuridos). El pedúnculo púbico puede presentar una rugosidad ovalada en su base, nombrada tuberosidad lateroventral (= lateroventral protuberances) (Prieto-Márquez, 2008), así como el foramen obturador expuesto lateralmente (Figura 10.5). Si este agujero no está completamente cerrado y se expone dorsoventralmente, se le denomina escotadura del obturador (= obturator gutter), rasgo único de los hadrosáuridos (Figura 10.1).
El ala prepúbica (= prepubic process, anterior pubic process, prepubic bar, cranial pubic process) es la estructura plana y de mayor extensión del pubis, surge cranealmente del cuerpo púbico (Figuras 10.1, 10.3). Esta región se divide en el cuello y la lámina púbicas (Brett-Surman y Wagner, 2007). El cuello púbico (= prepubic constriction) ocupa la constricción máxima del ala, entre el cuerpo y la lámina púbica (Figura 10.1, 10.3). Por el contrario, la lámina púbica (= pubic blade) es la expansión dorsoventral del ala, la parte más delgada y frágil de dicha región (Figura 10.1, 10.3).
El escapo púbico (= postpubic process, postpubic rod, pubic bar, caudal pubic process) es un proceso de forma acicular, dorsoventralmente plano, con el ápice ligeramente curvado en sentido dorsal, que surge de la región ventrocaudal del cuerpo púbico (Figuras 10.1–10.2). Del borde dorsomedial del escapo, nace una cresta que recorre toda la longitud del proceso, la cual forma la pared medial del surco obturador (Ramírez-Velasco et al., 2012; Figura 10.1).
De acuerdo con la forma de la lámina prepúbica, los pubis de los hadrosauroideos pueden ser agrupados en seis morfotipos (modificados de los caracteres de Prieto-Márquez, 2010; Xing et al., 2014a). El primero está ejemplificado en la mayoría de los hadrosauroideos basales (Figuras 10.4–10.5), donde su lámina es ovalada y corta, más larga que alta y con una expansión dorsal mayor a la ventral.
El segundo morfotipo corresponde a Tethyshadros, Gobihadros y la mayoría de los lambeosaurinos (Figuras 9.1, 10.6–10.7). En éste, la lámina tiene una forma cuadrangular con el borde ventral redondeado u obtuso, es más alta que larga y con una expansión mayor dorsal que ventral (Brett-Surman y Wagner, 2007; Prieto-Márquez, 2008; Xing et al., 2014a).
El tercer morfotipo (= morfotipo tres de Brett-Surman y Wagner, 2007) está presente en Hypacrosaurus y Saurolophus (Figuras 10.8–10.9). La lámina tiene una forma de hacha, más alta que larga y más expandida ventral que dorsalmente. Presenta dos ángulos bien definidos en la parte media de los bordes dorsal y ventral, el ángulo ventral es agudo y el dorsal obtuso.
El cuarto morfotipo está ejemplificado en los miembros de la tribu Brachylophosaurini (Figura 10.10). Su lámina es ovalada larga, con una mayor expansión ventral que dorsal y con los bordes formados por aristas rectas que dan lugar a ángulos redondeados, dirigidos dorsal, ventral y cranealmente (Prieto-Márquez, 2008).
El quinto morfotipo está tipificado por los Kritosaurini (Figura 10.11). Su lámina es rectangular, más larga que alta y fuertemente inclinada hacia ventral (Prieto-Márquez, 2008).
El sexto morfotipo se encuentra representado por los Edmontosaurini (Figura 10.1). Su lámina es semicircular y con una expansión ventral más profunda que la dorsal (Prieto-Márquez, 2008; Xing et al., 2014a).
Isquion. Es el elemento más largo de la cintura pélvica de los hadrosauroideos, distinguiéndose por su expansión craneal dorsoventral y una estructura larga caudalmente (Figuras 10.12–10.20). Comprende el cuerpo y el escapo isquiáticos (Brett-Surman y Wagner, 2007).
El cuerpo isquiático es una estructura en forma de una herradura plana, con su borde cóncavo que contribuye a la escotadura acetabular (Figuras 10.12–10.14). El cuerpo da lugar a dos proyecciones cuadrangulares o pedúnculos: el iliaco que se inclina dorsocaudalmente y el púbico que se dirige ventrocranealmente. En la mayoría de los hadrosauroideos se presenta una escotadura isquiopúbica (= obturator foramen, obturator fenestra) en el borde ventrocaudal del cuerpo, ésta se encuentra delimitada cranealmente por el pedúnculo y caudalmente por el proceso obturador (Figuras 10.15–10.20). Adicionalmente a estas estructuras, en algunos hadrosáuridos, existe un proceso ventral (Figuras 10.12, 10.14, 10.18–10.20) que se puede fusionar o no con el proceso obturador, para dar lugar al foramen isquiático (= obturator gutter) (Figuras 10.12–10.14).
El escapo isquiático (= ischial shaft, shaft) es una proyección larga, delgada y lateromedialmente plana (Figuras 10.12, 10.14). Su terminación caudal o ápice puede ser simple o estar expandida dorsoventralmente en forma de bota (bota isquiática), como ocurre en los hadrosauroideos basales y lambeosaurinos (Brett-Surman y Wagner, 2007). Usualmente, el eje longitudinal es recto, aunque puede estar arqueado ventralmente (e.g., Eolambia, Jinzhousaurus, Xuwulong, Probactrosaurus, Nanningosaurus y Nanyangosaurus) (McDonald et al., 2017; Figura 10.17) o dorsalmente (e.g., Hadrosaurus, hadrosaurio de Sabinas y Prosaurolophus) (Kirkland et al., 2006; Drysdale et al., 2019; Figura 10.18). Este rasgo, de acuerdo con McDonald et al. (2017), podría ser ontogenéticamente variable, como lo demuestra los ejemplares de Eolambia. La cara medial del escapo es plana y estriada cerca del extremo caudal, superficie sinficial con la cual se une con su contraparte (Figura 10.14).
En vista lateral, los isquiones de los hadrosauroideos pueden ser agrupados en tres morfotipos (Prieto-Márquez, 2008): el primero, representado en la mayoría de los hadrosauroideos basales (Figuras 10.15–10.17), tiene un pedúnculo iliaco con un borde caudal cóncavo, una escotadura isquiopúbica larga y abierta y, por lo general, tiene una bota isquiática semicircular (Brett-Surman y Wagner, 2007; Prieto-Márquez, 2008).
El segundo morfotipo está ejemplificado por Claosaurus, Tethyshadros, Hadrosaurus, Lophorhothon y todos los saurolofinos (Figuras 10.12, 10.18). El pedúnculo iliaco tiene bordes rectos, una escotadura pequeña y usualmente cerrada en individuos adultos, así como de una terminación caudal no expandida (Brett-Surman y Wagner, 2007; Prieto-Márquez, 2008). Como caso particular, Tethyshadros, Gryposaurus, el hadrosaurio de Sabinas, y los miembros de la tribu Edmontosaurini presentan escapos muy delgados y rectos (Brett-Surman y Wagner, 2007; Prieto-Márquez, 2008).
El tercer morfotipo lo tienen los lambeosaurinos (Figuras 9.1, 10.19–10.20), su isquion tiene un pedúnculo iliaco en forma de bota, por lo tanto, presenta un borde caudal fuertemente cóncavo. Su escotadura es pequeña y cerrada en formas adultas, con una bota isquiática bien definida, de borde craneal curvado ventrocranealmente (Brett-Surman y Wagner, 2007; Prieto-Márquez, 2008). Adicionalmente, el escapo suele ser más robusto que los anteriores morfotipos y su superficie lateral del cuerpo y parte del escapo muestra una depresión lateral longitudinal (Conti et al., 2020).
3.9. Miembro pélvico
Está compuesto por el fémur, los huesos de la pierna que incluyen a la tibia y fíbula, así como los huesos del pie como el tarso, metatarso y las falanges pedales.
Fémur. Es el elemento más largo y robusto del miembro pélvico de los hadrosauroideos (Horner et al., 2004). Como todo hueso largo, está conformado por un cuerpo y dos extremos (Figuras 11.1–11.8).
El cuerpo femoral tiene forma de prisma rectangular, recto en toda su longitud o con el extremo distal arqueado caudalmente (Figuras 11.1–11.8). En la mitad de su longitud, sobre su borde caudomedial, nace una cresta cuneiforme o deltada denominada cuarto trocánter (Figuras 11.1–11.3). La cara medial del cuarto trocánter está marcada por dos depresiones (impresiones musculares del caudofemoral y del puboisquiofemoral) (Maidment et al., 2014) y su cara lateral se encuentra marcada por una superficie convexa. En la superficie lateral del cuerpo femoral, algunos hadrosauroideos muestran la impresión muscular femorotibial (Figura 11.1) una depresión profunda y ovalada, ubicada a la altura del cuarto trocánter (Young, 1958, fig. 30; Dilkes, 2000; Brett-Surman y Wagner, 2007).
El extremo proximal muestra una proyección medial semiesférica denominada cabeza femoral, la cual está unida al resto del hueso por una corta constricción o cuello femoral (Figuras 11.1–11.5). En la cara lateral se encuentran dos salientes rugosas y planas, conocidas como trocánteres craneal y mayor (Figuras 11.1–11.4). El trocánter mayor se extiende desde la cara proximal hasta el primer cuarto proximal de la longitud total de la cara caudal. Tiene una forma cuadrangular u oblongada y su borde proximal está más elevado que la cabeza y el trocánter craneal. En cambio, el trocánter craneal (= lesser trochanter) es de forma linguliforme y estrecho, se halla distalmente al mayor y usualmente separado de él por una escotadura. En Pararhabdodon, la región proximal se encuentra arqueada en vista craneal/caudal (Serrano et al., 2021).
El extremo distal femoral está formado por dos eminencias circulares, el cóndilo medial y el lateral, con un diámetro mayor proyectado hacia caudal con respecto a su parte craneal (Figuras 11.1–11.4, 11.6). Ambos están separados cranealmente por un surco intercondilar (= anterior intercondylar groove, condylar canal, extensor tunnel, cranial depression) y caudalmente por una fosa poplítea (= posterior intercondylar groove, caudal depression) casi triangular y proyectada verticalmente (Figuras 11.2, 11.4, 11.6). En vista distal, las superficies articulares de los cóndilos son rugosas, con el cóndilo medial más ancho y robusto que el lateral (Figura 11.6). En la cara lateral del cóndilo lateral, se observa una excavación semicircular en su región caudal, denominada tróclea fibular (Figuras 11.1, 11.6, 11.7)
En los hadrosauroideos, se reconocen dos morfotipos femorales (Norman, 2002; Prieto-Márquez, 2008). En los hadrosauroidea basales, su cuerpo es enteramente recto o presenta una curvatura en su extremo distal, su surco intercondilar es abierto y poco profundo, así como un cuarto trocánter cuneiforme. Como rasgo adicional, en algunos hadrosauroideos, se puede observar el trocánter craneal fuertemente separado del trocánter mayor (Brownstein, 2020).
En la familia Hadrosauridae, el fémur es completamente recto, sus integrantes tienen el surco intercondilar más profundo y en formas adultas, éste se encuentra parcialmente cerrado en su extremo distal (Brett-Surman y Wagner, 2007; Figura 11.6). El cuarto trocánter tiene una forma deltada, el trocánter craneal está unido al trocánter mayor y muestra un pequeño surco que los separa. Respecto a la proporción del fémur longitud/ancho en vista craneal, la mayoría de los lambeosaurinos presentan formas más delgadas respecto a los hadrosauroideos basales, saurolofinos y los Parasaurolophini (Ramírez-Velasco et al., 2021a).
Tibia. Es larga, torsionada a lo largo de su eje, con una expansión craneocaudal en su extremo proximal y una lateromedial en su extremo distal. Tiene un cuerpo y dos extremos (Figuras 11.9–11.16).
El cuerpo tibial es un prisma triangular torsionado, cuenta con tres caras convexas y dos bordes conspicuos (Figuras 11.9–11.13). En vista caudal, el borde medial nace entre los dos maléolos distalmente y termina en la base del cóndilo medial (Figuras 11.9–11.10). En cambio, el borde lateral se origina del maléolo lateral y desaparece en los dos tercios proximales del cuerpo tibial (Figuras 11.10–11.11).
El extremo proximal está formado por la cresta cnemial, estructura prominente de forma triangular que abarca la mitad o más de la mitad de la longitud del hueso (Prieto-Márquez, 2008). Su superficie lateral es cóncava, la medial convexa y forma un ápice que se curva lateralmente (Figuras 11.10–11.12). En oposición a la cresta cnemial, se presentan dos masas óseas o cóndilos, uno lateral y otro medial (= inner condyle/outer condyle) (Figura 11.12). Ambos están separados por un área interarticular (= intercondylar groove) en forma de surco vertical.
El extremo distal tibial tiene forma deltada en vista craneo/caudal, debido a la presencia de los dos maléolos (medial y lateral) separados por una depresión denominada surco extensor tibial (Figura 11.11). Por esta estructura pasan los ligamentos extensores y se articula el proceso ascendente del astrágalo. El maléolo lateral (= external malleolus, outer malleolus) tiene una forma lanceolada en vista craneal (Figuras 11.9–11.11, 11.13), con una superficie caudal concavo-convexa (o tróclea del cartílago tibial) y una craneal plana (superficie articular fibular). El maléolo medial (= internal malleolus, inner malleolus) tiene forma cónica y es más pequeño que el lateral.
Las tibias de los hadrosauroideos se pueden separar en tres morfotipos (Godefroit et al., 1998; Prieto-Márquez, 2008; Conti et al., 2020). El primer morfotipo se reconoce en la mayoría de los hadrosauroideos basales como Bolong, Choyrodon, Probactrosaurus, Eolambia y Yunganglong (Figuras 11.14–11.15), en donde presenta una cresta cnemial corta que no sobrepasa el tercio proximal de la longitud del hueso, cuenta con una fuerte separación distal entre el maléolo lateral y el maléolo medial, así como un eje levemente arqueado en vista caudal (Brownstein, 2020).
Los morfotipos segundo y tercero comparten la presencia de una cresta cnemial larga extendida hasta la mitad de la longitud total de la tibia y un eje recto. Estos morfotipos se distinguen por la separación distal de los maléolos: en el morfotipo dos, observado en Edmontosaurini y Tsintaosaurini, presentan una fuerte separación de los maléolos (Figura 11.11), mientras en el morfotipo tres, están pobremente separados como ocurre en Saurolophus, Gryposaurus y en la mayoría de los lambeosaurinos (Figura 11.16).
Fíbula. Es un elemento largo, delgado y con extremos ligeramente expandidos (Horner et al., 2004). Como todo hueso largo presenta un cuerpo, una cabeza y un extremo distal (Figuras 11.17–11.26).
El cuerpo fibular es un semicilíndrico torsionado (Figuras 11.17–11.21). Cuenta con una superficie articular tibial que se distribuye a todo lo largo del cuerpo, localizada desde la cara medial proximal a la caudal, distalmente. En general, el eje del cuerpo es más amplio en el extremo proximal y se adelgaza progresivamente hacia al extremo distal.
La cabeza fibular o extremo proximal tiene forma espatulada en vista lateral, con un ápice anguloso hacia craneal (Figuras 11.17–11.20). Cuenta con una superficie convexa lateral y una cóncava medialmente, esta última para la articulación con la tibia. La cara proximal, ligeramente convexa y rugosa, es la superficie donde se articula el cóndilo lateral femoral (Figura 11.17).
El extremo o proceso distal (= fibular foot, distal end) representa la región expandida con forma triangular, que cuenta con una terminación redondeada cranealmente (Figuras 11.17–11.19, 11.21). Caudalmente, es una superficie plana y estriada con la que se articula el maléolo lateral de la tibia.
Entre los hadrosauroideos, se han identificado cinco morfotipos fibulares, contrariamente a los dos descritos por Brett-Surman y Wagner (2007). El primero se reconoce en Edmontosaurini y los hadrosauroideos basales, en donde la cabeza es poco expandida y el proceso distal pequeño (Figuras 11.17, 11.22). El segundo se conoce en los Brachylophosaurini, se caracterizan por tener una cabeza robusta más ancha que el extremo distal, y por presentar un proceso distal ligeramente expandido (Figura 11.23). El tercero se reconoce en Gryposaurus, cuyas fíbulas son delgadas, poseen una cabeza pequeña y un proceso fuertemente engrosado (Figura 11.24). El cuarto se presenta en la mayoría de los lambeosaurinos, es similar al morfotipo uno, con la diferencia de tener un proceso distal casi esférico (Figura 11.25). El quinto se observa en Parasaurolophus y Saurolophus, los cuales poseen fíbulas robustas, con una cabeza pequeña y un proceso distal con forma de bota en vista lateral (Figura 11.26).
Tarso. En los hadrosauroideos, incluyen tres elementos óseos dispuestos en dos hileras horizontales, una proximal y otra distal (Figura 12). Los tarsales proximales, están conformados por el astrágalo y el calcáneo, mientras que sólo se conoce el tarsal IV en la hilera distal.
El calcáneo (= fibulare) es un hueso corto y pequeño que se articula proximalmente con la fíbula, caudalmente con la tibia y medialmente con el astrágalo (Figuras 12.1–12.6). Es de forma piramidal truncado, en él se pueden distinguir una cresta tibiofibular y un cuerpo (Figuras 12.4–12.6). En vista proximal, el cuerpo tiene forma de semicírculo (Figura 12.5), lateralmente es deltado con un borde distal redondeado y forma parte del cóndilo lateral. La superficie lateral presenta una concavidad denominada depresión epicondilar lateral (Figura 12.4). En vista proximal, la cresta tibiofibular (= ascending process, posterolateral process) se proyecta lateromedialmente y separa dos superficies articulares cóncavas, la craneal para la fíbula y la caudal para la tibia (Figuras 12.5–12.6).
El astrágalo (= tibiale) es un hueso corto con forma de copa o acetabulado, separado por una cresta robusta en vista proximal (Figuras 12.7–12.9). Se articula proximalmente con la tibia y lateralmente con el calcáneo (Figuras 12.1–12.3). Se puede dividir en un cuerpo y un proceso ascendente. La cara proximal del cuerpo presenta dos superficies articulares tibiales, la medial más amplia y la lateral de forma rectangular y de menor tamaño (Figura 12.8). En vista distal, el cuerpo forma dos superficies articulares rugosas, el cóndilo medial y el cóndilo lateral, separados por una escotadura intercondilar (Figura 12.2). El proceso ascendente es una estructura deltada en vista craneal, con una excavación llamada depresión intercondilar (Figura 11.9), proyectada craneocaudalmente en forma de una cresta baja, la cual separa las dos superficies articulares proximales (Figuras 12.7–12.9). En vista proximal, su extremo craneal es más ancho y alto que su extremo caudal (Figura 12.8).
El tarsal IV es una estructura discoidal de superficie rugosa, cóncava proximalmente y convexa distalmente (Brett-Surman y Wagner, 2007; Prieto-Márquez, 2007; Zheng et al., 2011; Figura 12.16).
De los elementos que conforman el tarso de los hadrosauroideos, sólo el astrágalo presenta cambios morfológicos notables (Prieto-Márquez, 2008; Brett-Surman y Wagner, 2007). En la mayoría de los hadrosauroideos basales y saurolofinos, la cara craneal es tan amplia como la tibia (Figuras 12.1, 12.9–12.12). En cambio, en los lambeosaurinos es más pequeña y sólo cubre la parte central de la tibia (Figuras 12.3, 12.13–12.15). Además, la depresión intercondilar presenta cambios en su tamaño, pues es grande y amplia en la mayoría de los hadrosauroideos, y relativamente pequeña en Edmontosaurus y Parasaurolophus (Figuras 12.9, 12.13).
Metatarso. En los hadrosauroideos, el metatarso está conformado por los metatarsianos II, III y IV (Figuras 13.1–13.3). De acuerdo con Horner et al. (2004), a lo largo de la historia de los Ornithopoda, el metatarsiano I se reduce hasta quedar como estructura vestigial en los iguanodóntidos y finalmente, se pierde en los Hadrosauroidea. Como todo hueso largo, se compone de un cuerpo y dos extremos (Figuras 13.4–13.15).
El metatarsiano II tiene un cuerpo semicilíndrico, comprimido mediolateralmente y medialmente arqueado (Figuras 13.4–13.7). La superficie lateral es de plana a ligeramente cóncava y la medial es convexa. Cerca del extremo distal, en su borde laterodorsal, presenta una pequeña cresta denominada lámina mediodorsal (= lateral knob, mediodorsal flange) (Prieto-Márquez, 2010; Figuras 13.4–13.5). En vista proximal, su extremo es elíptico y de superficie cóncava. Cuenta con dos crestas, una dorsal y otra plantar (Figura 13.6). El extremo distal o tróclea metatarsal tiene una forma rectangular distalmente y es más alto que ancho (Figura 13.7).
El metatarsiano III es el más largo y robusto. El cuerpo es cilíndrico recto, su extremo proximal es expandido dorsoplantarmente y el distal es más deprimido (Figuras 13.8–13.11). En su terminación proximal presenta una cresta dorsal y plantar pequeñas y una superficie cóncava profunda para la articulación con el metatarsiano II medialmente (Figura 13.8, 13.10). La tróclea tiene forma rectangular, más ancha que alta (Figura 13.11).
El metatarsiano IV tiene un cuerpo semicilíndrico, lateralmente arqueado (Figuras 13.12–13.15). La cara medial es ligeramente cóncava y la lateral convexa. Los extremos están expandidos dorsoplantarmente (Figuras 13.14–13.15). Cercano a su terminación proximal, en su cara medial, presenta una superficie rugosa ovalada, la tuberosidad medial (= craneomedial knob) (Prieto-Márquez, 2008), estructura distintiva de este metatarsiano (Figura 13.12–13.13). La tróclea en vista distal tiene forma cuadrangular a rectangular y es más alta que ancha (Figura 13.15).
La forma y estructura de los metatarsianos no presentan cambios notorios en los hadrosauroideos. Quizás la única excepción es la mayor robustez de éstos en los hadrosauroideos basales, comparada con los hadrosáuridos (Prieto-Márquez, 2001; 2010; McDonald et al., 2012). Como rasgo único, en Augustynolophus presenta un metatarsiano III con una cresta dorsal de forma cónica (Prieto-Márquez y Wagner, 2013, fig. 5).
Falanges pedales. Se pueden clasificar en falanges proximales (II-1, III-1 y IV-1), medias (II-2; III-2 a III-3; y IV-2 a IV-4) y distales o falanges ungueales (II-3, III-4 y IV-5). Usualmente, son largas y en el extremo distal, presentan depresiones circulares en cada costado, denominadas fóveas del ligamento colateral. La fórmula falángica pedal 0/3/4/5/0 es constante en este grupo (Horner et al., 2004). Al ser huesos largos, están provistos de un cuerpo y dos extremos (Figuras 13.16–13.26).
Las falanges proximales varían en forma, desde prismas rectangulares asimétricos (Figura 13.16, 13.18) o simétricos (Figura 13.17), a prismas triangulares (Figura 13.19). Estas formas dependen de su posición en el pie, son simétricas en el dígito III y asimétricas en el II y IV. La forma prismática triangular es única en el dígito IV de los hadrosauroideos basales.
Las falanges medias pueden variar de forma cuadrangular (Figuras 13.20–13.21) a discoidal (Figura 13.22). Curiosamente, la forma cuadrangular de la falange media es típica de los hadrosauroideos basales y de los saurolofinos, ubicada en el dígito II (Figuras 13.1, 13.3).
Los ungueales pedales de los hadrosauroideos son estructuras deprimidas, desde lanceoladas a forma de medialuna (Figuras 13.23–13.26). Dependiendo del dígito, éstas pueden ser asimétricas, si se ubican en el dígito II y IV, o simétricas en el dígito III (Figuras 13.1–13.3). Las ungueales de los hadrosauroideos basales son lanceoladas, largas y con un surco lateral (Figura 13.23). En cambio, en los hadrosáuridos, las ungueales tienen forma de medialuna en vista dorsopalmar y carecen del surco lateral (Figuras 13.24-13.26) (Prieto-Márquez, 2008). Como caso especial, las ungueales de los Brachylophosaurini y Lapampasaurus presentan una quilla plantar (= axial keel) en el ungueal III (Figuras 13.25–13.26).
4. Conclusiones
Contrario a las ideas generales previas, las cuales señalaban que el postcráneo de los hadrosauroideos carecía de elementos diagnósticos para la identificación y descripción de nuevas especies, esta guía exhibe numerosos caracteres útiles para tales fines, como previamente lo han señalado Brett-Surman y Wagner (2007) y posteriormente, Prieto-Márquez (2010). Cada hueso, de forma individual, presenta un conjunto de características que permiten discernir la categoría taxonómica, incluyendo rasgos distintivos a nivel de especie, incluso si se tienen elementos aislados, incompletos o fragmentados. Estos rasgos se concentran principalmente en los extremos de los huesos largos (e.g., húmero, radio, ulna, metacarpo, fémur, tibia, fíbula, metatarso y falanges) y planos (e.g., escápula, coracoides, placa esternal, ilion, pubis e isquion), así como en la región dorsal (arco vertebral) de la columna vertebral.
Este proyecto surgió ante la necesidad de una nueva recopilación bibliográfica sobre la anatomía de los hadrosauroideos y es la primera en su tipo en habla hispana. El trabajo ofrece una guía detallada, ilustrada y actualizada para jóvenes estudiantes o investigadores que necesiten describir e identificar material completo o fragmentado de estos dinosaurios. En esta ocasión, sólo se centró en los elementos óseos postcraneanos y subsiguientemente, ofreceremos un trabajo complementario que incluya los elementos craneanos. Por último, esperamos que esta guía sea útil para los investigadores y estudiantes de este grupo.
Agradecimientos
El presente trabajo surge como parte de la tesis doctoral del primer autor, dentro del programa de Doctorado del Posgrado de Ciencias Biológicas de la UNAM (5199) en el Instituto de Geología de la UNAM. Se agradece a R. Hernández Rivera, M. Montellano Ballesteros y L.M. Ochoa Ochoa por sus comentarios, que ayudaron a mejorar la calidad del manuscrito. A los curadores de las colecciones V. Romero Mayen (IGM), J.M. Padilla Gutiérrez (CPC), V.M. Escalante Hernández (CPC), M.C. Aguillón-Martínez (CPC), F. Aguilar (CIC), H.G. Porras Múzquiz (MUZ) y R. Guajardo (PASAC) por la ayuda en la revisión del material de sus respectivas colecciones. A los revisores anónimos y a los editores de la revista por sus atinadas sugerencias que mejoraron el manuscrito final. Agradecemos particularmente a la editora en jefe la Dra. Ana Bertha Villaseñor, a la editora técnica la Mtra. S. Ramos Amézquita y al formador el Mtro. L.F. Álvarez Sánchez. Este proyecto fue financiado por la UNAM, a través de los proyectos DGAPA-PAPIIT IN IN207314 e IN110920, y por la Beca de Doctorado del CONACYT (303851) otorgada al primer autor.
Referencias
Allen, H., 2009, Guide to common vertebrate fossils from the Cretaceous of Alberta: Calgary, Alberta, Alberta Paleontological Society, 234 pp.
Álvarez del Villar, J., Álvarez, T., Álvarez-Castañeda, S.T., 2007, Diccionario de anatomía comparada de vertebrados: D.F., México, Instituto Politécnico Nacional, 252 pp.
Baumel, J.J., Witmer, L.M., 1993, Osteologia, en Baumel, J.J., King, A., Lucas Breazile, A., Evans, H. (eds.), Handbook of avian anatomy: nomina anatomica avium: Cambridge, Massachusetts, Publication of the nuttal ornithological club, 45–136.
Bolotsky, Y.L., Godefroit, P., 2004, A new hadrosaurine dinosaur from the Late Cretaceous of Far Eastern Russia: Journal of Vertebrate Paleontology, 24(2), 351–365, doi: 10.1671/1110
Bolotsky, Y.L., Kurzanov, S.K., 1991, The hadrosaurs of the Amur Region, en Moiseyenko, V.G. (ed.), Geology of the Pacific Ocean Border: Blagoveschensk, Rusia, Amur KNII, far Eastern Branch of the Russian Academy of Sciences, 94–103.
Borinder, N.H., Poropat, S.F., Campione, N.E., Wigren, T., Kear, B. P., 2021, Postcranial osteology of the basally branching hadrosauroid dinosaur Tanius sinensis from the Upper Cretaceous Wangshi Group of Shandong, China: Journal of Vertebrate Paleontology, e1914642, doi: 10.1080/02724634.2021.1914642
Boulenger, G.A., 1881, Sur l´arc pelvien chez les dinosauriens de Bernissart [On the pelvic arch in the dinosaurs of Bernissart]: Bulletins de L'Académie royale de Belgique, 3, 1–11.
Brett-Surman, M.K., 1979, Phylogeny and paleobiogeography of hadrosaurian dinosaurs: Nature 277(5697), 560–562.
Brett-Surman, M.K., 1989, A revision of the Hadrosauridae (Reptilia: Ornithischia) and their evolution during the Campanian and Maastrichtian: Washington, George Washington University, tesis doctoral, 192 pp.
Brett-Surman, M.K., Wagner, J.R., 2007, Discussion of character analysis of the appendicular anatomy in Campanian and Maastrichtian North America hadrosaurids-variation and ontogeny, en Carpenter, K. (ed.), Horns and beaks: ceratopsian and ornithopod dinosaurs: Bloomington, Indiana University Press, 135–169.
Brown, B., 1910, The Cretaceous Ojo Alamo Beds of New Mexico with description of the new dinosaur genus Kritosaurus: Bulletin of the American Museum of Natural History, 28, 267–274.
Brown, B., 1912, A crested dinosaur from the Edmonton Cretaceous: Bulletin of the American Museum of Natural History, 31, 131–136.
Brown, B., 1913, The skeleton of Saurolophus, a crested duck-billed dinosaur from the Edmonton Cretaceous: Bulletin of the American Museum of Natural History, 32, 387–393.
Brown, B., 1914, Corythosaurus casuarius, a new crested dinosaur from the Belly River Cretaceous, with provisional classification of the family Trachodontidae: Bulletin of the American Museum of Natural History, 33, 559–565.
Brown, B., 1916, A new crested trachodont dinosaur Prosaurolophus maximus: Bulletin of the American Museum of Natural History, 35, 701–708.
Brownstein, C.D. 2020, Osteology and phylogeny of small-bodied hadrosauromorphs from an end-Cretaceous marine assemblage: Zoological Journal of the Linnean Society, 191(1), 180–200, doi: 10.1093/zoolinnean/zlaa085
Brownstein, C.D., Bissell, I. 2020, An elongate hadrosaurid forelimb with biological traces informs the biogeography of the Lambeosaurinae: Journal of Paleontology, 95(2), 367–375, doi: 10.1017/jpa.2020.83
Buffetaut, E., Suteethorn, V., 2011, A new iguanodontian dinosaur from the Khok Kruat Formation (Early Cretaceous, Aptian) of northeastern Thailand: Annales de Paléontologie, 97(1–2), 51–62, doi: 10.1016/j.annpal.2011.08.001
Butler, R.J., Barrett, P.M., Gower, D.J., 2012, Reassessment of the evidence for postcranial skeletal pneumaticity in Triassic archosaurs, and the early evolution of the Avian Respiratory System: PLoS ONE, 7 (3), e34094, doi: 10.1371/journal.pone.0034094
Campione, N., 2014, Postcranial anatomy of Edmontosaurus regalis (Hadrosauridae) from the Horseshoe Canyon Formation, Alberta, Canada, en Eberth, D.A., Evans, D.C. (eds.), Hadrosaurs: Bloomington, Indiana University Press, 208–244, https://hdl.handle.net/1959.11/27865
Carpenter, K., Ishida, Y., 2010, Early and “middle” Cretaceous iguanodonts in time and space: Journal of Iberian Geology, 36(2), 145–164, doi: 10.5209/rev_JIGE.2010.v36.n2.3
Casanovas-Cladellas, M.L., Santafé-Llopis, J.V., Isidro-Llorens, A., 1993, Pararhabdodon isonensis n. gen. n. sp. (Dinosauria). Estudio mofológico, radio-tomográfico y consideraciones biomecánicas: Paleontologia i Evolució, 26–27, 121–131.
Conti, S., Vila, B., Sellés, A.G., Galobart, A., Benton, M.J., Prieto-Márquez, A., 2020, The oldest lambeosaurine dinosaur from Europe: insights into the arrival of Tsintaosaurini: Cretaceous Research, 107, 104286, doi: 10.1016/j.cretres.2019.104286
Coria, R.A., González Riga, B., Casadio, S., 2012, Un nuevo hadrosáurido (Dinosauria, Ornithopoda) de la Formación Allen, provincia de La Pampa, Argentina: Ameghiniana, 49(4), 552–572, doi: 10.5710/AMGH.9.4.2012.487
Cruzado-Caballero, P., Powell, J.E., 2017, Bonapartesaurus rionegrensis, a new hadrosaurine dinosaur from South America: implications for phylogenetic and biogeographic relations with North America: Journal of Vertebrate Paleontology, 37(2), e1289381, doi: 10.1080/02724634.2017.1289381
Cruzado-Caballero, P., Pereda-Suberbiola, X., Ruiz-Omeñaca, J.I., 2010, Blasisaurus canudoi gen. et sp. nov., a new lambeosaurine dinosaur (Hadrosauridae) from the Latest Cretaceous of Arén (Huesca, Spain): Canadian Journal of Earth Sciences, 47(12), 1507–1517, doi: 10.1139/E10-081
Dalla Vecchia, F.M., 2009, Tethyshadros insularis, a new hadrosauroid dinosaur (Ornithischia) from the Upper Cretaceous of Italy: Journal of Vertebrate Paleontology, 29(4), 1100–1116, doi: 10.1671/039.029.0428
Dalla Vecchia, F.M., 2020, The unusual tail of Tethyshadros insularis (Dinosauria, Hadrosauroidea) from the Adriatic Island of the European archipelago: Revista Italiana di Paleontologia e Stratigrafia, 126(3), 583–628, doi: 10.13130/2039-4942/14075
Dilkes, D.W., 2000, Appendicular myology of the hadrosaurian dinosaur Maiasaura peeblesorum from the Late Cretaceous (Campanian) of Montana: Transactions of the Royal Society of Edinburgh, Earth Sciences, 90, 87–125.
Dorian, A.F., 1988, Elsevier´s encyclopedic dictionary of medicine part B anatomy: Amsterdam, Elsevier, 601 pp.
Dorland, W.A.N., 2012, Dorland´s illustrated medical dictionary, Treintadoceava edición: EUA, Saunders, 2147 pp.
Drysdale, E.T., Therrien, F., Zelenitsky, D.K., Weishampel, D.B., Evans, D.C., 2019, Description of juvenile specimens of Prosaurolophus maximus (Hadrosauridae: Saurolophinae) from the Upper Cretaceous Bearpaw Formation of southern Alberta, Canada, reveals ontogenetic changes in crest morphology: Journal of Vertebrate Paleontology, 38(6), e1547310-2, doi: 10.1080/02724634.2018.1547310
Frandson, R.D., Wilke, W.L., Fails, A.D., 2009, Anatomy and physiology of farm animals, Séptima edición: EUA, Wiley-Blackwell publishing, 512 pp.
Freedman, E.A., 2015, Evolution and ontogeny of hadrosaurs (Dinosauria: Ornithischia) in the Judith River Formation (Late Cretaceous: Campanian) of Northcentral Montana; Montana, Montana State University, tesis doctoral, 477 pp.
Freedman, E.A.F., Horner, J.R., 2015, A New brachylophosaurin hadrosaur (Dinosauria: Ornithischia) with an intermediate nasal crest from the Campanian Judith River Formation of Northcentral Montana: PLoS ONE, 10(11), e0141304, doi: 10.1371/journal.pone.0141304
Gates, T.A., Sampson, S.D., 2007, A new species of Gryposaurus (Dinosauria: Hadrosauridae) from the late Campanian Kaiparowits Formation, southern Utah, USA: Zoological Journal of the Linnean Society, 151(2), 351–376, doi: 10.1111/j.1096-3642.2007.00349.x
Gates, T.A., Scheetz, R., 2014, A new saurolophine hadrosaurid (Dinosauria: Ornithopoda) from the Campanian of Utah, North America: Journal of Systematic Palaeontology, 13(8), 711-725, doi: 10.1080/14772019.2014.950614
Gates, T.A., Horner, J.R., Hanna, R.R., Nelson, C.R., 2011, New unadorned hadrosaurine hadrosaurid (Dinosauria, Ornithopoda) from the Campanian of North America: Journal of Vertebrate Paleontology, 31(4), 798–811, doi: 10.1080/02724634.2011.577854
Gates, T.A., Jinnah, Z., Levitt, C., Getty, M.A., 2014, New hadrosaurid specimens from the lower-middle Campanian Wahweap Formation of Utah, en Eberth, D.A., Evans, D.C. (eds.), Hadrosaurs: Bloomington, Indiana University Press, 156–173.
Gates, T.A., Sampson, S., Delgado de Jesús, C.R., Zanno, L.E., Eberth, D.A., Hernández-Rivera, R., Aguillón-Martínez, M.C., Kirkland, J.I., 2007, Velafrons coahuilensis, a new lambeosaurine hadrosaurid (Dinosauria: Ornithopoda) from the Late Campanian Cerro del Pueblo Formation, Coahuila, Mexico: Journal of Vertebrate Paleontology, 27(4), 917–930, doi: 10.1671/0272-4634(2007)27%5B917:VCANLH%5D2.0.CO;2
Gates, T.A., Tsogtbaatar, K., Zanno, L.E., Chinzoring, T., Watabe, M., 2018, A new iguanodontian (Dinosauria: Ornithopoda) from the Early Cretaceous of Mongolia: PeerJ, 6, e5300, doi: 10.7717/peerj.5300
Gilmore, C.W., 1933, On the dinosaurian fauna of the Iren Dabasu Formation: Bulletin of the American Museum of Natural History, 67, 23–78, http://hdl.handle.net/2246/355.
Gilmore, C.W., 1935, On the reptilia of the Kirttland Formation of New Mexico, with descriptions of new species of fossil turtles: Proceedings of the United States National Museum, 83(2978), 159–188, doi: 10.5479/si.00963801.83-2978.159
Godefroit, P., Dong, Z.M., Bultynck, P., Li, H., Feng, L., 1998, New Bactrosaurus (Dinosauria: Hadrosauroidea) material from Iren Dabasu (Inner Mongolia, P.R. China): Sciences de la Terre, 68, 3–70.
Godefroit, P., Zan, S., Jin, L., 2000, Charonosaurus jiayinensis n.g., n.sp.,a lambeosaurine dinosaur from the Late Maastrichtian of northeastern China: Comptes Rendus de l'Academie des Sciences Series IIA Earth and Planetary Science, 330(12), 875–882, doi: 10.1016/S1251-8050(00)00214-7
Godefroit, P., Zan, S., Jin, L., 2001, The Maastrichtian (Late Cretaceous) lambeosaurine dinosaur Charonosaurus jiayinensis from north-eastern China: Bulletin de l´institut royal des sciences naturelles de Belgique Science de la Terre, 71, 119–168.
Godefroit, P., Bolotsky, Y., Alifanov, V., 2003, A remarkable hollow-crested hadrosaur from Russia: an Asian origin for lambeosaurines: Comptes Rendus Palevol, 2(2), 143–151, doi: 10.1016/S1631-0683(03)00017-4
Godefroit, P., Bolotsky, Y., Van Itterbeeck, J., 2004, The lambeosaurine dinosaur Amurosaurus riabinini, from the Maastrichtian of Far Eastern Russia: Acta Palaeontologica Polonica, 49(4), 585–618.
Godefroit, P., Li, H., Shang, C.Y., 2005, A new primitive hadrosauroid dinosaur from the Early Cretaceous of Inner Mongolia (P.R. China): Comptes Rendus Palevol, 4(8), 697–705, doi: 10.1016/j.crpv.2005.07.004
Godefroit, P., Hai, S., Yu, T., Lauters, P., 2008, New hadrosaurid dinosaurs from the uppermost Cretaceous of north−eastern China: Acta Palaeontologica Polonica, 53(1), 47–74, doi: 10.4202/app.2008.0103
Godefroit, P., Bolotsky, Y.L., Lauters, P., 2012a, A new saurolophine dinosaur from the latest Cretaceous of Far Eastern Russia: PLoS ONE, 7(5), e36849, doi: 10.1371/journal.pone.0036849
Godefroit, P., Bolotsky, Y.L., Bolotsky, I.Y., 2012b, Osteology and relationships of Olorotitan arharensis, a hollow−crested hadrosaurid dinosaur from the latest Cretaceous of Far Eastern Russia: Acta Palaeontologica Polonica, 57(3), 527–560, doi: 10.4202/app.2011.0051
Godefroit, P., Escuillié, F., Bolotsky, Y.L., Lauters, P., 2012c, A new basal hadrosauroid dinosaur from the Upper Cretaceous of Kazakhstan, en Godefroit, P. (ed.), Bernissart Dinosaurs and Early Cretaceous Terrestrial Ecosystems: EUA, Indiana University Press, 335–358.
Guenther, F.M., 2009, Influence of sequence heterochrony on hadrosaurid dinosaur postcranial development: The Anatomical record, 292, 1427–1441, doi: 10.1002/ar.20988
Harris, J.D., 2004, Confusing dinosaurs with mammals: tetrapod phylogenetics and anatomical terminology in the world of homology: The Anatomical record part A, 281(2), 1240–1246, doi: 10.1002/ar.a.20078
Head, J.J., 1998, A new species of basal hadrosaurid (Dinosauria, Ornithopoda) from the Cenomanian of Texas: Journal of Vertebrate Paleontology 18(4), 718–738.
Hendrickx, C., Mateus, O., 2014, Torvosaurus gurneyi n. sp., the largest terrestrial predator from Europe, and a proposed terminology of the maxilla anatomy in nonavian theropods: PLoS ONE, 9(3): e88905, doi: 10.1371/journal.pone.0088905
Hohloch, A., 2013, On the tracks of Brachylophosaurus: simulation-based examinations on the locomotion of hadrosaurs: Alemania, Universidad de Tubinga, tesis doctoral, 192 pp.
Horner, J.R., 1992, Cranial morphology of Prosaurolophus (Ornithischia: Hadrosauridae) with descriptions of two new hadrosaurid species and an evaluation of hadrosaurid phylogenetic relationships: Museum of Rockies Occasional Paper, 2, 1–119.
Horner, J.R., Currie, P.J., 1994, Embryonic and neonatal morphology and ontogeny of a new species of Hypacrosaurus (Ornithischia, Lambeosauridae) from Montana and Alberta, en Carpenter, K., Hirsch, K.F., Horner, J.R. (eds.), Dinosaur eggs and babies: Cambridge, Cambridge University Press, 312–336.
Horner, J.R., Makela, R., 1979, Nest of juveniles provides evidence of family structure among dinosaurs: Nature, 282, 296–298, doi: 10.1038/282296a0
Horner, J.R., Weishampel, D.B., Forster, C.A., 2004, Hadrosauridae, en Weishampel, D.B., Dodson, P., Osmólska, H. (eds.), The dinosauria, second edition: Berkeley, University of California Press, 438–463, doi: 10.1525/california/9780520242098.003.0023
Hu, C.C., 1973, A new hadrosaur from the Cretaceous of Chucheng, Shantung: Acta Geologica Sinica, 2, 179–206.
Hu, C., Cheng, Z., Pang, Q., Fang, X., 2001, Shantungosaurus giganteus: Beijing, China, Geological Publishing House, 206 pp.
Hunt, A.P., Lucas, S.G., 1993, Cretaceous vertebrates of New Mexico: New Mexico Museum of Natural History and Science Bulletin, 2, 77–91.
Hutchinson, J.R., 2001, The evolution of pelvic osteology and soft tissues on the line to extant birds (Neornithes): Zoological Journal of Linneo Society, 131(2), 123–168, doi: 10.1111/j.1096-3642.2001.tb01313.x
Kirkland, J.I., 1998, A new hadrosaurid from the upper Cedar Mountain Formation (Albian-Cenomanian: Cretaceous) of eastern Utah – the oldest known hadrosaurid (lambeosaurine?): New Mexico Museum of Natural History and Science Bulletin, 14, 283–295.
Kirkland, J.I., Hernández-Rivera, R., Gates, T., Paul, G.S., Nesbitt, S., Serrano-Brañas, C.I., García-de la Garza, P.J., 2006, Large hadrosaurine dinosaurs from the Latest Campanian of Coahuila, Mexico: New Mexico Museum of Natural History and Sciences Bulletin, 35, 299–315.
Kobayashi, Y., Nishimura, T., Takasaki, R., Chiba, K., Fiorillo, A.R., Tanaka, K., Tsogtbaatar, C., Sato, T., Sakurai, K., 2019, A new hadrosaurine (Dinosauria: Hadrosauridae) from the marine deposits of the Late Cretaceous Hakobuchi Formation, Yezo Group, Japan: Scientific Reports, 9 (12389), doi: 10.1038/s41598-019-48607-1
Kobayashi, Y., Takasaki, R., Kubota, K., Fiorillo, A.R., 2021, A new basal hadrosaurid (Dinosauria: Ornithischia) from the latest Cretaceous Kita-ama Formation in Japan implies the origin of hadrosaurids: Scientific Reports, 11 (8547), doi: 10.1038/s41598-021-87719-5
Lambe, L.M., 1914, On Gryposaurus notabilis, a new genus and species of trachodont dinosaur from the Belly River Formation of Alberta, with a description of the skull of Chasmosaurus belli: The Ottawa Naturalist, 27, 145–155.
Lambe, L.M., 1917, A new genus and species of crestless hadrosaur from the Edmonton Formation of Alberta: The Ottawa Naturalist, 31, 65–73.
Lambe, L.M., 1920. The hadrosaur Edmontosaurus from the Upper Cretaceous of Alberta, Geological Survey of Canada, Memoir, 120: Ottawa, T. Mulvey, Printer, , 79 pp, doi: 10.4095/101655
Langstone, W.Jr., 1960, The vertebrate fauna of the Selma Formation of Alabama. Part VI. The dinosaurs: Fieldiana, Geology Memoirs, 3(6), 317–361, doi: 10.5962/bhl.title.5344
Lehman, T.M., Wick, S.L., Wagner, J.R., 2016, Hadrosaurian dinosaurs from the Maastrichtian Javelina Formation Big Bend National Park, Texas: Journal of Paleontology, 90(2), 333–356, doi: 10.1017/jpa.2016.48
Leidy, J., 1858, Hadrosaurus foulkii, a new saurian from the Cretaceous of New Jersey, related to Iguanodon: Proceedings of the Academy of Natural Sciences of Philadelphia, 10, 213–218.
Longrich, N.R., Pereda-Suberbiola, X., Pyron, R.A., Jalil, N.E., 2020, The first duckbill dinosaur (Hadrosauridae: Lambeosaurinae) from Africa and the role of oceanic dispersal in dinosaur biogeography: Cretaceous Research, 120, 104678, doi: 10.1016/j.cretres.2020.104678
Lü, J.C., 1997, A new Iguanodontidae (Probactrosaurus mazongshanensis sp. nov.) from Mazongshan area, Gansu Province, en Dong, Z. (ed.), Sino-Japanese Silk Road Dinosaur Expedition: Beijing, China, China Ocean Press, 27–47.
Lull, R.S., Wrigth, N.E., 1942, Hadrosaurian dinosaurs of North America, en Aldrich, H.R. (ed.), Geological Society of America, Special Papers Number 40: Baltimore, Waverly Press, 242 pp., doi: 10.1130/SPE40-p1
Main, D.J., Noto, C.R., Weishampel, D.B., 2014, Postcranial anatomy of a basal hadrosauroid (Dinosauria: Ornithopoda) from the Cretaceous (Cenomanian) Woodbine Formation of North Texas, en Eberth, D.A., Evans, D.C. (eds.), Hadrosaurs: Bloomington, Indiana University Press, 77–95.
Maidment, S.C.R., Bates, K.T., Barrett, P.M., 2014, Three-dimensional computational modeling of pelvic locomotor muscle moment arms in Edmontosaurus (Dinosauria, Hadrosauridae) and comparisons with Other Archosaurs, en Eberth, D.A., Evans, D.C. (eds.), Hadrosaurs: Bloomington, Indiana University Press, 433–448.
Marsh, O.C., 1872, Notice on a new species of Hadrosaurus: American Journal of Science, 3(16), 301.
Marsh, O.C., 1892, Notice of new reptiles from the Laramie Formation: American Journal of Science, 43(257), 449–453, doi: 10.2475/ajs.s3-43.257.449
Maryanska, T., Osmólska, H., 1981, First lambeosaurine dinosaur from the Nemegt Formation, Upper Cretaceous, Mongolia: Acta Palaeontologica Polonica, 26(3–4), 243–255.
McDonald, A.T., Wolfe, D.G., Kirkland, J.I., 2010, A new basal hadrosauroid (Dinosauria: Ornithopoda) from the Turonian of New Mexico: Journal of Vertebrate Paleontology, 30(3), 799–812, doi: 10.1080/02724631003763516
McDonald, A.T., Bird, J., Kirkland, J.I., Dodson, P., 2012, Osteology of the basal hadrosauroid Eolambia caroljonesa (Dinosauria: Ornithopoda) from Cedar Mountain Formation of Utah: PLoS ONE, 7(10), e45712, doi: 10.1371/journal.pone.0045712
McDonald, A.T., Maidment, S.C.R., Barrett, P.M., You, H.I., Dodson, P., 2014, Osteology of the basal hadrosauroid Equijubus normani (Dinosauria, Ornithopoda) from the Early Cretaceous of China, en Eberth, D.A., Evans, D.C. (eds.), Hadrosaurs: Bloomington, Indiana University Press, 44–72.
McDonald, A.T., Gates, T.A., Zanno, L.E., Makovicky, P.J., 2017, Anatomy, taphonomy, and phylogenetic implications of a new specimen of Eolambia caroljonesa (Dinosauria: Ornithopoda) from the Cedar Mountain Formation, Utah, USA: PLoS ONE, 12(5), e0176896, doi: 10.1371/journal.pone.0176896
McDonald, A.T., Wolfe, D.G., Freedman Fowler, E.A., Gates, T.A., 2021, A new brachylophosaurin (Dinosauria: Hadrosauridae) from the Upper Cretaceous Menefee Formation of New Mexico: PeerJ, 9, e11084, doi: 10.7717/peerj.11084
Mo, J., Zhao, Z., Wang, W., Xu, X., 2007, The first hadrosaurid dinosaur from southern China: Acta Geologica Sinica, 81(4), 550–554, doi: 10.1111/j.1755-6724.2007.tb00978.x
Morris, W.J., 1981, A new species of hadrosaurian dinosaur from the Upper Cretaceous of Baja California—?Lambeosaurus laticaudus: Journal of Paleontology, 55(2), 453–462.
Nagao, T., 1936, Nipponosaurus sachalinensis – A new genus and species of trachodont dinosaur from Japanese Saghalien: Journal of the Faculty of Science, Hokkaido Imperial University Series 4, Geology and mineralogy, 3(2), 185–220, http://hdl.handle.net/2115/35768
Nagao, T., 1983, On the limb-bones of Nipponosaurus sachalinensis Nagao, a Japanese hadrosaurian dinosaur: The Zoological Society of Japan, 17(3–4), 311–319.
Nopcsa, B.F., 1900, Dinosaurierreste aus Siebenbürgen (Schädel von Limnosaurus transsylvanicus nov. gen. et spec.) [Dinosaur remains from Transylvania (skull of Limnosaurus transsylvanicus nov. gen. et spec.)]: Denkschriften der Kaiserlichen Akademie der Wissenschaften, Mathematisch-Naturwissenschaftliche Classe, 68, 555–591.
Norman, D.B., 1998, On Asian ornithopods (Dinosauria: Ornithischia). 3. A new species of iguanodontid dinosaur: Zoological Journal of the Linnean Society, 122(1–2), 291–348, doi: 10.1111/j.1096-3642.1998.tb02533.x
Norman, D.B., 2002, On asian ornithopods (Dinosauria: Ornithischia). 4. Probactrosaurus Rozhdestvensky, 1966: Zoological Journal of the Linnean Society, 136, 113–144.
Organ, C.L., Adams, J., 2005, The histology of ossified tendon in dinosaurs: Journal of Vertebrate Paleontology, 25(3), 602–613, doi: 10.1671/0272-4634(2005)025%5B0602:THOOTI%5D2.0.CO;2
Ostrom, J.H., 1963, Parasaurolophus cyrtocristatus, a crested hadrosaurian dinosaur from New Mexico: Fieldiana, Geology, 14(8), 143–168, doi: 10.5962/bhl.title.5353
Parks, W.A., 1922, Parasaurolophus walkeri, a new genus and species of crested trachodont dinosaur: University of Toronto Studies: Geological Series, 13, 1–32.
Parks, W.A., 1923, Corythosaurus intermedius, a new species of trachodont dinosaur: University of Toronto Studies: Geological Series, 15, 1–57.
Pereda-Suberbiola, X., Canudo, J.I., Cruzado-Caballero, P., Barco, J.L., López-Martínez, N., Oms, O., Ruiz-Omeñaca, J.I., 2009, The last hadrosaurid dinosaurs of Europe: A new lambeosaurine from the Uppermost Cretaceous of Aren (Huesca, Spain): Comptes Rendus Palevol, 8(6), 559–572, doi: 10.1016/j.crpv.2009.05.002
Prieto-Márquez, A., 2001, Osteology and variation of Brachylophosaurus canadensis (Dinosauria, Hadrosauridae) from the Upper Cretaceous Judith River Formation of Montana: Bozeman, Montana, Montana State University, tesis de maestría, 390 pp.
Prieto-Márquez, A., 2007, Postcranial osteology of the hadrosaurid Brachylophosaurus canadensis from the Late Cretaceous of Montana, en Carpenter, K. (ed.), Horns and beaks: ceratopsian and ornithopod dinosaurs: Bloomington, Indiana University Press, 91–115.
Prieto-Márquez, A., 2008, Phylogeny and historical biogeography of hadrosaurid dinosaurs: Florida, Florida State University College of Arts and Sciences, tesis de doctorado, 861 pp.
Prieto-Márquez, A., 2010, The braincase and skull roof of Gryposaurus notabilis (Dinosauria, Hadrosauridae), with a taxonomic revisión of the genus: Journal of Vertebrate Paleontology, 30(3), 838–854, doi: 10.1080/02724631003762971
Prieto-Márquez, A., 2011, Revised diagnoses of Hadrosaurus foulkii Leidy, 1858 (the type genus and species of Hadrosauridae Cope, 1869) and Claosaurus agilis Marsh, 1872 (Dinosauria: Ornithopoda) from the Late Cretaceous of North America: Zootaxa, 2765(1), 61–68, doi: 10.11646/zootaxa.2765.1.6
Prieto-Márquez, A., 2012, The skull and apendicular skeleton of Gryposaurus latidens, a saurolophine hadrosaurid (Dinosauria: Ornithopoda) from the early Campanian (Cretaceous) of Montana, USA: Canadian Journal of Earth Science, 49(3), 510–532, doi: 10.1139/e11-069
Prieto-Márquez, A., 2014, A juvenile Edmontosaurus from the Late Maastrichtian (Cretaceous) of North America: implications for ontogeny and phylogenetic inference in saurolophine dinosaurs: Cretaceous Research, 50, 282–303, doi: org/10.1016/j.cretres.2014.05.003
Prieto-Márquez, A., Carrera Farias, M.Á., 2021, A new late-surviving early diverging Ibero-Armorican duck-billed dinosaur and the role of the Late Cretaceous European Archipelago in hadrosauroid biogeography: Acta Palaeontologica Polonica, 66(2), 425–435, doi: 10.4202/app.00821.2020
Prieto-Márquez, A., Gutarra, S., 2016, The “duck-billed” dinosaurs of Careless Creek (Upper Cretaceous of Montana, USA), with comments on hadrosaurid ontogeny: Journal of Paleontology, 90(1), 133–146, doi: 10.1017/jpa.2016.42
Prieto-Márquez, A., Norell, M., 2010, Anatomy and relationships of Gilmoreosaurus mongoliensis (Dinosauria, Hadrosauroidea) from the late Cretaceous of Central Asia: American Museum Novitates, 3694, 1–49, doi: 10.1206/3694.2
Prieto-Márquez, A., Salinas, G.C., 2010, A re-evaluation of Secernosaurus koerneri and Kritosaurus australis (Dinosauria, Hadrosauridae) from the Late Cretaceous of Argentina: Journal of Vertebrate Paleontology, 30(3), 813–837, doi: 10.1080/02724631003763508
Prieto-Márquez, A., Serrano Brañas, C.I., 2012, Latirhinus uitstlani, a ‘broad-nosed’ saurolophine hadrosaurid (Dinosauria, Ornithopoda) from the late Campanian (Cretaceous) of northern Mexico: Historical Biology, 24(6), 607–619, doi: 10.1080/08912963.2012.671311
Prieto-Márquez, A., Wagner, J.R., 2013, A new species of saurolophine hadrosaurid from the Late Cretaceous of the Pacific coast of North America: Acta Palaeontologica Polonica, 58(2), 255–268, doi: 10.4202/app.2011.0049
Prieto-Márquez, A., Chiappe, L.M., Joshi, S.H., 2012, The lambeosaurine dinosaur Magnapaulia laticaudus from the Late Cretaceous of Baja California, Northwestern Mexico: PLoS ONE, 7(6), e38207, doi: 10.1371/journal.pone.0038207
Prieto-Márquez, A., Dalla Vecchia, F.M., Gaete, R., Galobart, A., 2013, Diversity, relationships, and biogeography of the lambeosaurine dinosaurs from the European Archipelago, with description of the new aralosaurin Canardia garonnensis: PLoS ONE, 8(7), e69835, doi: 10.1371/journal.pone.0069835
Prieto-Márquez, A., Erickson, G.M., Ebersole, J.A., 2016, A primitive hadrosaurid from southeastern North America and the origin and early evolution of 'duck-billed' dinosaurs: Journal of Vertebrate Paleontology, 36(2), e1054495, doi: 10.1080/02724634.2015.1054495
Prieto-Márquez, A., Wagner, J.R., Lehman, T., 2019a, An unusual “shovel-billed” dinosaur with trophic specializations from the early Campanian of Trans-Pecos Texas, and the ancestral hadrosaurian crest: Journal of Systematic Palaeontology, 18(6), 1–38, doi: 10.1080/14772019.1625078
Prieto-Márquez, A., Fondevilla, V., Sellés, A. G., Wagner, J. R., Galobart, À., 2019b, Adynomosaurus arcanus, a new lambeosaurine dinosaur from the Late Cretaceous Ibero-Armorican Island of the European Archipelago: Cretaceous Research, 96, 19–37, doi: 10.1016/j.cretres.2018.12.002
Ramírez-Velasco, A.A., Benammi, M., Prieto-Márquez, A., Alvarado Ortega, J., Hernández-Rivera, R., 2012, Huehuecanauhtlus tiquichensis, a new hadrosauroid dinosaur (Ornithischia: Ornithopoda) from the Santonian (Late Cretaceous) of Michoacán, Mexico: Canadian Journal of Earth Sciences, 49(2), 379–395, doi: 10.1139/e11-062
Ramírez-Velasco, Á.A., Espinosa-Arrubarrena, L., Alvarado-Ortega, J., 2021a, Review of the taxonomic affinities of Latirhinus uitstlani, an emblematic Mexican hadrosaurid: Journal of South American Earth Sciences, 110, 103391, doi: 10.1016/j.jsames.2021.103391
Ramírez-Velasco, A.A., Aguilar, F.J., Hernández-Rivera, R., Gudiño Maussán, J.L., Lara Rodríguez, M., Alvarado-Ortega, J., 2021b, Tlatolophus galorum, gen. et sp. nov., a parasaurolophini dinosaur from the upper Campanian of the Cerro del Pueblo Formation, Coahuila, northern Mexico: Cretaceous Research, 126, 104884, doi: 10.1016/j.cretres.2021.104884
Riabinin, A.N., 1938, Some results of the studies of the Upper Cretaceous dinosaurian fauna from the vicinity of the station Sary-Agach, South Kazakhstan: Problems of Paleontology, 4, 125–135.
Rozhdestvensky, A.K., 1952, A new representative of the duck-billed dinosaurs from the Upper Cretaceous deposits of Mongolia: Dokladi Akademii Nauk S.S.S.R. 86, 405–408.
Rozhdestvensky, A.K., 1966, Novyye iguanodonty iz Tsentral'noy Azii. Filogeneticheskiye i taksonomicheskiye v zaimootnosheniya pozdnikh Iguanodontidae i rannikh Hadrosauridae [New iguanodonts from Central Asia. Phylogenetic and taxonomic interrelationships of late Iguanodontidae and early Hadrosauridae]: Paleontologicheskii Zhurnal, 3, 103–116.
Rozhdestvensky, A.K., 1968, Gadrozavry Kazakhstana [Hadrosaurs of Kazakhstan]., en Tatarinov, L.P, et al. (eds.), [Upper Paleozoic and Mesozoic Amphibians and Reptiles]: Moscow, Akademia Nauk SSSR, 97–141.
Senter, P., 2012, Forearm orientation in Hadrosauridae (Dinosauria: Ornithopoda) and implications for museum mounts: Palaeontologia Electronica, 15(3), 10, doi: 10.26879/330
Sereno, P.C., 1986, Phylogeny of the bird-hipped dinosaurs: National Geographic Research, 2(2), 234–256.
Serrano, J.F., Sellés, A.G., Vila, B., Galobart, Á., Prieto-Márquez, A., 2021, The osteohistology of new remains of Pararhabdodon isonensis sheds light into the life history and paleoecology of this enigmatic European lambeosaurine dinosaur: Cretaceous Research, 118, 104677, doi: 10.1016/j.cretres.2020.104677
Shibata, M., Azuma, Y., 2015, New basal hadrosauroid (Dinosauria: Ornithopoda) from the Lower Cretaceous Kitadani Formation, Fukui, central Japan: Zootaxa, 3914(4), 421–440, doi: 10.11646/zootaxa.3914.4.3
Shibata, M., Jintasakul, P., Azuma, 2011, A new iguanodontian dinosaur from the Lower Cretaceous Khok Kruat Formation, Nakhon Ratchasima in Northeastern Thailand: Acta Geologica Sinica, 85(5), 969–976, doi: 10.1111/j.1755-6724.2011.00530.x
Shibata, M., Jintasakul, P., Azuma, Y., You, H.L., 2015, A new basal hadrosauroid dinosaur from the Lower Cretaceous Khok Kruat Formation in Nakhon Ratchasima province, Northeastern Thailand: PLoS ONE, 10(12), e0145904, doi: 10.1371/journal.pone.0145904
Stefanic, C.M., Nesbitt, S.J., 2019, The evolution and role of the hyposphene-hypantrum articulation in Archosauria: phylogeny, size and/or mechanics?: Royal Society Open Science, 6(10), 190258, doi: 10.1098/rsos.190258
Sternberg, C.M., 1935, Hooded hadrosaurs of the Belly River Series of the Upper Cretaceous: Canada Department of Mines Bulletin, Geological Series, 52(77), 1–37.
Sternberg, C.M., 1953, A new hadrosaur from the Oldman Formation of Alberta: Discussion of nomenclatura: Canadian Department of Resource Development Bulletin, 128, 1–12.
Sues, H.D., Averianov, A., 2009, A new basal hadrosauroid dinosaur from the Late Cretaceous of Uzbekistan and the early radiation of duck-billed dinosaurs: Proceedings of the Royal Society B: Biological Sciences, 276(1667), 2549–2555, doi: 10.1098/rspb.2009.0229
Takasaki, R., Fiorillo, A.R., Tykoski, R.S., Kobayashi, Y., 2020, Re-examination of the cranial osteology of the Arctic Alaskan hadrosaurine with implications for its taxonomic status: PLoS ONE, 15(5), e0232410, doi: 10.1371/journal.pone.0232410
Taylor, D.G., Lucas S.G., 2018, A late Cretaceous (Campanian) hadrosaur sacrum from the Cape Sebastian sandstone, Curry County, Oregon: Fossil Record 6, New Mexico Museum of Natural History and Science Bulletin, 79, 695–702.
Tsogtbaatar, K., Weishampel, D.B., Evans, D.C., Watabe, M., 2014. A new hadrosauroid (Plesiohadros djadokhtaensis) from the Late Cretaceous Djadokhtan fauna of Southern Mongolia, en Eberth, D.A., Evans, D.C. (eds.), Hadrosaurs: Bloomington, Indiana University Press, 108–135.
Tsogtbaatar, K., Weishampel, D.B., Evans, D.C., Watabe, M., 2019, A new hadrosauroid (Dinosauria: Ornithopoda) from the Late Cretaceous Baynshire Formation of the Gobi Desert (Mongolia): PLoS ONE, 14(4), e0208480, doi: 10.1371/journal.pone.0208480
Wagner, J.R., Lehman, T.M., 2009, An enigmatic new lambeosaurine hadrosaur (Reptilia: Dinosauria) from the Upper Shale Mmember of the campanian Aguja Formation of trans-pecos Texas: Journal of Vertebrate Paleontology, 29(2), 605–611, doi: 10.1671/039.029.0208
Wang, X.L., Xu, X., 2001, A new iguanodontid (Jinzhousaurus yangi gen. et sp. nov.) from the Yixian Formation of western Liaoning, China: Chinese Science Bulletin, 46(19), 1669–1672, doi: 10.1007/BF02900633
Wang, X., Pan, R., Butler, R.J., Barrett, P.M., 2011, The postcranial skeleton of the iguanodontian ornithopod Jinzhousaurus yangi from the Lower Cretaceous Yixian Formation of western Liaoning, China: Earth and Enviromental Science Transactions of the Royal Society of Edinburgh, 101(2), 135–159, doi: 10.1017/S1755691010009266
Wang, R.F., You, H.L., Xu, S.C., Wang, S.Z., Yi, J., Xie, L.J., Jia, L., Li, Y.X., 2013, A new hadrosauroid dinosaur from the early Late Cretaceous of Shanxi Province, China: PLoS ONE, 8(10), e77058, 1–12, doi: 10.1371/journal.pone.0077058
Wang, R.F., You, H.L., Wang, S.Z., Xu, S.C., Yi, J., Xie, L.J., Jia, L., Xing, H., 2015, A second hadrosauroid dinosaur from the early Late Cretaceous of Zuoyun, Shanxi province, China: Historical Biology, 29(1), 17–24, doi: 10.1080/08912963.2015.1118688
Weishampel, D.B., Horner, J.R., 1990, Hadrosauridae, en Weishampel, D.B., Dodson, P., Osmólska, H. (eds), The Dinosauria: Berkeley, University of California Press, 534–551.
White, T.D., Black, M.T., Folkens, P.A., 2012, Human Osteology, Tercera edición: California, Oxford, Academic Press, 662 pp.
Wilson, J.A., 1999, A nomenclatura for vertebral laminae in sauropods and other saurischian dinosaurs: Journal of Vertebrate Paleontology, 19(4), 639–653, doi: 10.1080/02724634.1999.10011178
Wiman, C., 1929, Die Kreide-Dinosaurier aus Shantung: Palaeontologia Sinica, Series C, 6, 1–67.
Wiman, C., 1931, Parasaurolophus tubicen n. sp. aus der Kreide in New Mexico [Parasaurolophus tubicen n. sp. from the Cretaceous in New Mexico]: Nova Acta Regiae Societatis Scientarum Upsaliensis, Series IV, 7(5), 3–11.
Wu, W., Godefroit, P., 2012, Anatomy and relationships of Bolong yixianensis, an Early Cretaceous iguanodontoid dinosaaur from Western Liaoning, China, en Godefroit, P. (ed), Bernissart Dinosaurs and Early Cretaceous Terrestrial Ecosystems: Indiana University Press, 293–333.
Wu, W., Godefroit, P., Hu, D., 2010, Bolong yixianensis gen. et sp. nov.: A new Iguanodontoid dinosaur from the Yixian Formation of Western Liaoning, China: Geology and Resources, 19(2), 127–133.
Xing, H., Zhao, X., Wang, K., Li, D., Chen, S., Mallon, J.C., Zhang, Y., Xu, X., 2014a, Comparative Osteology and phylogenetic relationship of Edmontosaurus and Shantungosaurus (Dinosauria: Hadrosauridae) from the Upper Cretaceous of North America and East Asia: Acta Geological Sinica, 88(6), 1801–1840, doi: 10.1111/1755-6724.12334
Xing, H., Wang, D., Han, F., Sullivan, C., Ma, Q., He, Y., Hone, D.W.E., Yan, R., Du, F., Xu, X., 2014b, A new basal hadrosauroid dinosaur (Dinosauria: Ornithopoda) with transitional features from the Late Cretaceous of Henan Province, China: PLoS ONE, 9(6), e98821, doi: 10.137/journal.pone.0098821
Xu, X., Zhao, X.J., Lü, J.C., Huang, W.B., Li, Z.L., Dong, Z.M., 2000, A new iguanodontian from Sangping Formation of Neixiang, Henan and its stratigraphical implication: Vertebrata PalAsiatica, 38(3), 176–191.
Xu, S.C., You, H.L., Wang, J.W., Wang, S.Z., Yi, J., Yia, L., 2016, A new hadrosauroid dinosaur from the Late Cretaceous of Tianzhen, Shanxi Province, China: Vertebrata PalAsiatica, 54, 67–78, doi: 10.19615/j.cnki.1000-3118.2016.01.005
You, H.L., Li, D.Q., 2009, A new basal hadrosauriform dinosaur (Ornithischia: Iguanodontia) from the Early Cretaceous of northwestern China: Canadian Journal of Earth Sciences, 46(12), 949–957, doi: 10.1139/E09-067
You, H., Li, D., Liu, W., 2011, A new hadrosauriform dinosaur from the Early Cretaceous of Gansu Province, China: Acta Geologica Sinica, 85(1), 51–57, doi: 10.1111/j.1755-6724.2011.00377.x
You, H.L., Ji, Q., Li, J.L., Li, Y.X., 2003a, A new hadrosauroid dinosaur from the mid-Cretaceous of Liaoning, China: Acta Geologica Sinica, 77, 148–154, doi: 10.1111/j.1755-6724.2003.tb00557.x
You, H.L., Luo, Z.X., Shubin, N.H., Witmer, L.M., Tang, Z.L., Tang, F., 2003b, The earliest-known duck-billed dinosaur from deposits of late Early Cretaceous age in northwest China and hadrosaur evolution: Cretaceous Research, 24(3), 347–355, doi: 10.1016/S0195-6671(03)00048-X
Young, C.C., 1958, The dinosaurian remains of Laiyang, Shantung: Palaeontologia Sinica, New series C, 42(16), 1–138.
Zhang, J.L., Wang, X.L., Wang, Q., Jiang, S.X., Cheng, X., Li, N., Qiu, R., 2017, A new saurolophine hadrosaurid (Dinosauria: Ornithopoda) from the Upper Cretaceous of Shandong, China: Anais da Academia Brasileira de Ciências, 91(2), doi: 10.1590/0001-376520160920
Zheng, R., Farke, A.A., Kim, G.S., 2011, A photographic atlas of the pes from a hadrosaurine hadrosaurid dinosaur: PalArch´s Journal of Vertebrate Paleontology, 8(7), 1–8.
Zheng, W., Jin, X., Shibata, M., Azuma, Y., 2014. An early juvenile specimen of Bolong yixianenesis (Ornithopoda: Iguanodontia) from the Lower Cretaceous of Ningcheng County, Nei Mongol, China: Historical Biology, 26(2), 236–251, doi: 10.1080/08912963.2013.809347
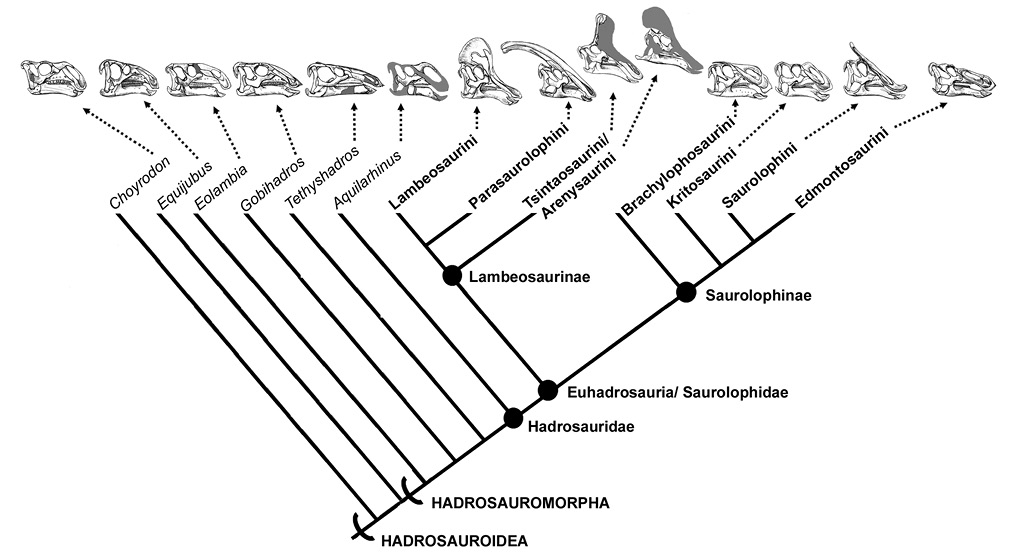
Figura 1. Relación filogenética simplificada de los hadrosauroideos.
Tabla 1. Lista de Hadrosauroidea válidos
|
Taxón / referencia |
Localidad |
Edad |
|
Acristavus gagslarsoni Gates, Horner, Hanna y Nelson, 2011 |
Fm. Two Medicine (Montana), EUA; Fm. Wahweap (Utah), EUA |
Campaniano |
|
Adelolophus hutchisoni Gates, Jinnah, Levitt y Getty, 2014 |
Fm. Wahweap (Utah), EUA |
Campaniano |
|
Adynomosaurus arcanus Prieto-Márquez, Fondevilla, Sellés, Wagner y Galobart, 2019b |
Fm. Conques (Leida), España |
Maastrichtiano tardío |
|
Augustynolophus morrisi (Prieto-Márquez y Wagner, 2013) |
Fm. Moreno (California), EUA |
Maastrichtiano tardío |
|
Ajnabia odysseus Longrich, Pereda Suberbiola, Pyron y Jalil, 2020 |
(Marruecos), África |
Maastrichtiano tardío |
|
Altirhinus kurzanovi Norman, 1998 |
Fm. Khuren Dukh (Gobi), Mongolia |
Albiano medio a tardío |
|
Amurosaurus riabinini Bolotsky y Kurzanov, 1991 |
Fm. Udurchukan (Amur), Rusia |
Maastrichtiano medio a tardío |
|
Angulomastacator daviesi Wagner y Lehman, 2009 |
Fm. Aguja (Texas), EUA |
Campaniano tardío |
|
Aquilarhinus palimentus Prieto-Márquez, Wagner y Lehman, 2019a |
Fm. Aguja (Texas), EUA |
Campaniano temprano |
|
Aralosaurus tuberiferus Rozhdestvensky, 1968 |
Fm. Bostobe, Kazakhstán |
Santoniano tardío a Campaniano temprano |
|
Arenysaurus ardevoli Pereda-Superbiola, Canudo, Cruzado-Caballero, Barco, López-Martínez, Oms y Ruiz-Omeñaca, 2009 |
Fm. Tremp (Pirineos), España |
Cretácico tardío |
|
Bactrosaurus johnsoni Gilmore, 1933 |
Fm. Iren Dabasu (Mongolia Interior), China |
Campaniano tardío a Maastrichtiano temprano |
|
Barsboldia sicinskii Maryanska y Osmólska, 1981 |
Fm. Nemegt (Nemegt), Mongolia |
Maastrichtiano medio |
|
Batyrosaurus rozhdestvenskyi Godefroit, Escuillié, Bolotsky y Lauters, 2012 |
(Kyzylorda), Kazakhstán |
Santoniano a Campaniano |
|
Blasisaurus canudoi Cruzado-Caballero, Pereda-Suberbiola y Ruiz-Omeñaca, 2010 |
Fm. Arén (Huesca), España |
Maastrichtiano tardío |
|
Bolong yixianensis Wu, Godefroit y Hu, 2010 |
Fm. Yixian (Liaoning), China |
Barremiano |
|
Bonapartesaurus rionegrensis Cruzado-Caballero y Powell, 2017 |
Fm. Allen (Río Negro), Argentina |
Campaniano tardío a Maastrichtiano temprano |
|
Brachylophosaurus canadensis Sternberg, 1953 |
Fm. Judith River (Montana), EUA; Fm. Oldman (Alberta), Canadá |
Campaniano tardío |
|
Canardia garonnensis Prieto-Márquez, Dalla Vecchia, Gaete y Galobart, 2013 |
Fm. Marnes d´Auzas (Petites-Pyrénées), Francia |
Maastrichtiano tardío |
|
Charonosaurus jiayinensis Godefroit, Zan y Jin, 2000 |
Fm. Yuliangze (Heilongjang), China |
Maastrichtiano tardío |
|
Choyrodon barsboldi Gates, Tsogtbaatar, Zanno, Chinzoring y Watabe, 2018 |
Fm. Khuren Dukh, Mongolia |
Albiano medio a tardío |
|
Claosaurus agilis (Marsh, 1872) |
Fm. Niobrara Chalk (Kansas), EUA |
Coniaciano tardío |
|
Corythosaurus casuarius Brown, 1914 |
Fm. Dinosaur Park (Alberta), Canadá |
Campaniano tardío |
|
Corythosaurus intermedius Parks, 1923 |
Fm. Dinosaur Park (Alberta), Canadá |
Campaniano tardío |
|
Datonglong tianzhenensis Xu, You, Wang J, Wang S, Yi y Yia, 2016 |
Fm. Huiquanpu (Sjanxi), China |
Cretácico tardío |
|
Edmontosaurus annectens Marsh 1892 |
Fm. Hell Creek (Montana, Dakota del Sur), EUA; Fm. Lance (Dakota del sur y Wyoming), EUA; Fm. Scollard (Alberta) Canadá; Fm. Frenchman (Saskatchewan), Canadá |
Maastrichtiano tardío |
|
Edmontosaurus regalis Lambe, 1917 |
Fm. Hell Creek (Montana, Dakota del sur), EUA; Fm. Lance (Wyoming), EUA; Fm. Laramie (Colorado), EUA; Fm. Prince Creek, (Alaska), EUA; Fm. Scollard, St. Mary, River y Horeshoe Canyon (Alberta), Canadá |
Maastrichtiano temprano a tardío |
|
Eolambia caroljonesa Kirkland, 1998 |
Fm. Cedar Mountain (Utah), EUA |
Cenomaniano temprano |
|
Eotrachodon orientalis Prieto-Márquez, Erickson y Ebersole, 2016 |
Fm. Mooreville Chalk (Alabama), EUA |
Santoniano tardío |
|
Equijubus normani You, Luo, Shubin, Witmer, Tang Z y Tang F, 2003b |
Gpo. Xinminbao (Gansu), China |
Barremiano a Albiano |
|
Fylax thyrakolasus Prieto-Márquez y Carrera Farias, 2021 |
Fm. Figuerola (Lleida), España |
Maastrichtiano tardío |
|
Gilmoreosaurus mongoliensis (Gilmore, 1933) |
Fm. Iren Dabasu (Mongolia Interior), China |
Campaniano tardío a Maastrichtiano temprano |
|
Gobihadros mongoliensis Tsogtbaatar, Weishampel, Evans y Watabe, 2019 |
Fm. Bayunshire (Desierto del Gobi), Mongolia |
Cenomaniano al Santoniano |
|
Gongpoquansaurus mazongshanensis (Lü, 1997) |
Fm. Zhonggou (Gansu), China |
Albiano |
|
Gryposaurus alsatei? Lehman, Wick y Wagner, 2016 |
Fm. Javelina (Texas), EUA |
Maastrichtiano medio o tardío |
|
Gryposaurus latidens Horner, 1992 |
Fm. Two Medicine (Montana), EUA |
Santoniano tardío a Campaniano temprano |
|
Gryposaurus notabilis Lambe, 1914 |
Fm. Dinosaur Park (Alberta), Canadá |
Campaniano tardío |
|
Gryposaurus monumentensis Gates y Sampson, 2007 |
Fm. Kaiparowits (Utah), EUA |
Campaniano tardío |
|
Hadrosaurus foulkii Leidy, 1858 |
Fm. Woodbury (New Jersey), EUA |
Campaniano |
|
Huehuecanauhtlus tiquichensis Ramírez-Velasco, Benammi, Prieto-Márquez, Alvarado-Ortega y Hernández-Rivera, 2012 |
(Michoacán), México |
Santoniano tardío |
|
Hypacrosaurus altispinus Brown, 1912 |
Fm. Horseshoe Canyon (Alberta), Canadá |
Maastrichtiano temprano |
|
Hypacrosaurus stebingeri Horner y Currie, 1994 |
Fm. Two Medicine (Montana), EUA; Fm. Oldman (Alberta), Canadá |
Campaniano medio al tardío |
|
Jaxartosaurus aralensis Riabinin, 1938 |
Fm. Syuksyuk, Kazakhstán |
Santoniano |
|
Jeyawati rugoculus McDonald, Wolfe y Kirkland 2010 |
Fm. Moreno Hill (New Mexico), EUA |
Turoniano medio |
|
Jintasaurus meniscus You y Li, 2009 |
Gpo. Xinminpu (Gansu), China |
Aptiano tardío |
|
Jinzhousaurus yangi Wang y Xu, 2001 |
Fm. Yixian (Liaoning), China |
Aptiano temprano |
|
Kamuysaurus japonicus Kobayashi, Nishimura, Takasaki, Chiba, Fiorillo, Tanaka, Tsogtbaatar, Sato y Sakurai, 2019 |
Fm. Hakobuchi (Hokkaido), Japón |
Maastrichtiano tardío |
|
Kazaklambia convincens (Rhozdestvensky, 1968) |
Dabrazinskaya Svita, Uzbekistán |
Santoniano |
|
Kerberosaurus manakini Bolotsky y Godefroit, 2004 |
Fm. Tsagayan (Amur), Rusia; Fm. Udurchukan (Amur), Rusia |
Maastrichtiano temprano |
|
Koshisaurus katsuyama Shibata y Azuma, 2015 |
Fm. Kitadani (Fukui), Japón |
Barremiano a Aptiano temprano |
|
Kritosaurus navajovius Brown, 1910 |
Fm. Upper Kirtland (New Mexico), EUA; Fm. Cerro del Pueblo (Coahuila), México |
Campaniano tardío |
|
Kritosaurus horneri (Hunt y Lucas, 1993) |
Fm. Lower Kirtland (New Mexico), EUA |
Campaniano tardío |
|
Laiyangosaurus youngi Zhang, Wang X, Wang Q, Jiang, Cheng, Li y Qiu, 2017 |
Fm. Jingangkou (Shandong), China |
Cretácico tardío |
|
Lambeosaurus clavinitialis Sternberg, 1935 |
Fm. Dinosaur Park (Alberta), Canadá |
Campaniano tardío |
|
Lambeosaurus lambei Parks, 1923 |
Fm. Dinosaur Park (Alberta), Canadá |
Campaniano tardío |
|
Lambeosaurus magnicristatus Sternberg, 1935 |
Fm. Dinosaur Park (Alberta), Canadá |
Campaniano tardío |
|
Lapampasaurus cholinoi Coria, González Riga y Casadio, 2012 |
Fm. Allen (La Pampa), Argentina |
Campaniano tardío a Maastrichtiano temprano |
|
Latirhinus uitstlani Prieto-Márquez y Serrano-Brañas, 2012 |
Fm. Cerro del Pueblo (Coahuila), México |
Campaniano tardío |
|
Levnesovia transoxiana Sues y Averianov, 2009 |
Fm. Bissekty (Navoi Viloyat), Uzbekistán |
Turoniano medio a tardío |
|
Lophorhothon atopus Langstone, 1960 |
Fm. Mooreville Chalk (Alabama), EUA |
Campaniano temprano |
|
Magnapaulia laticaudus (Morris, 1981) |
Fm. El Gallo (Baja California), México |
Campaniano tardío |
|
Maiasaura peeblesorum Horner y Makela, 1979 |
Fm. Two Medicine (Montana), EUA |
Campaniano medio al tardío |
|
Naashoibitosaurus ostromi Hunt y Lucas, 1993 |
Fm. Kirtland (New Mexico), EUA |
Campaniano tardío |
|
Nanningosaurus dashiensis Mo, Zhao, Wang y Xu, 2007 |
Red beds (Guangxi), China |
Cretácico tardío |
|
Nanyangosaurus zhugeii Xu, Zhao, Lü, Huang, Li y Dong, 2000 |
Fm. Sangping (Henan), China |
Turoniano al Campaniano |
|
Nipponosaurus sachalinensis Nagao, 1936 |
Gpo. Yezo superior (Sakhaklin), Rusia |
Santoniano tardío a Campaniano temprano |
|
Olorotitan ararhensis Godefroit, Bolotsky y Alifanov, 2003 |
Fm. Kundur (Amur), Rusia |
Maastrichtiano medio a tardío |
|
Ornatops incantatus McDonald, Wolfe, Freedman Fowler y Gates, 2021 |
Fm. Menefee (New Mexico), EUA |
Campaniano medio |
|
Pararhabdodon isonensis Casanovas-Caldellas, Santafé-Llopis e Isidro-Llorens, 1993 |
Fm. Tremp (Catalonia), España |
Maastrichtiano tardío |
|
Parasaurolophus cyrtocristatus Ostrom, 1963 |
Fm. Kaiparowits (Utah), EUA; Fm. Fruitland (Nuevo México), EUA |
Campaniano tardío |
|
Parasaurolophus tubicen Wiman, 1931 |
Fm. Kirtland (Nuevo México), EUA |
Campaniano tardío |
|
Parasaurolophus walkeri Parks, 1922 |
Fm. Dinosaur Park (Alberta), Canadá |
Campaniano tardío |
|
Penelopognathus weishampeli Godefroit, Li y Shang, 2005 |
Fm. Bayan Gobi (Mongolia Interior), China |
Albiano |
|
Plesiohadros djadokhtaensis Tsogtbaatar, Weishampel, Evans y Watabe, 2014 |
Fm. Djadokhta (Gobi), Mongolia |
Campaniano |
|
Probactrosaurus gobiensis Rozhdestvensky, 1966 |
Fm. Dashuigou (Mongolia Interior), China |
Albiano |
|
Probrachylophosaurus bergei Freedman y Horner, 2015 |
Fm. Judith River (Montana), EUA |
Campaniano tardío |
|
Prosaurolophus maximus Brown 1916 |
Fm. Dinosaur Park (Alberta), Canadá |
Campaniano tardío |
|
Protohadros byrdi Head, 1998 |
Fm. Woodbine (Texas), EUA |
Cenomaniano medio |
|
Ratchasimasaurus suranareae Shibata, Jintasakul y Azuma, 2011 |
Fm. Khok Kruat (Ratchasima), Tailandia |
Aptiano |
|
Rhinorex condrupus Gates y Scheetz, 2014 |
Fm. Neslen (Utah), EUA |
Campaniano tardío a Maastrichtiano temprano |
|
Sahaliyania elunchunorum Godefroit, Hai, Yu y Lauters, 2008 |
Fm. Yuliangze (Heilongjang), China |
¿Maastrichtiano tardío? |
|
Saurolophus angustirostris Rozhestvensky, 1952 |
Fm. Nemegt (Nemegt), Mongolia |
Maastrichtiano medio |
|
Saurolophus osborni Brown, 1912 |
Fm. Horseshoe Canyon (Alberta), Canadá |
Maastrichtiano temprano |
|
Secernosaurus koerneri Brett-Surman, 1979 |
Fm. Bajo Barreal (Chubut), Fm. Los Alamitos (Patagonia), Argentina |
Campaniano tardío al Maastrichtiano temprano |
|
Shantungosaurus giganteus Hu, 1973 |
Fm. Xingezhuang (Heilogjang), China; Fm. Hongtuya (Shandong), China |
Campaniano medio al tardío |
|
Shuangmiaosaurus gilmorei You, Ji, Li y Li, 2003a |
Fm. Sunjiawan (Liaoning), China |
Cretácico tardío |
|
Siamodon nimngami Buffetaut y Suteethorn, 2011 |
Fm. Khok Kruat (Ratchasima), Tailandia |
Aptiano |
|
Sirindhorna khoratensis Shibata, Jintasakul, Azuma y You, 2015 |
Fm. Khok Kruat (Ratchasima), Tailandia |
Aptiano |
|
Tanius sinensis Wiman, 1929 |
Fm. Jiangjun (Shandong), China |
Campaniano temprano |
|
Telmatosaurus transsylvanicus Nopcsa, 1900 |
Fm. Sampetru (Judetul Hunedoara), Rumania |
Maastrichtiano temprano |
|
Tethyshadros insularis Dalla Vecchia, 2009 |
Fm. Liburnian (Trieste), Italia |
Campaniano tardío a Maastrichtiano temprano |
|
Tlatolophus galorum Ramírez-Velasco, Aguilar, Hernández-Rivera, Gudiño Maussán, Lara Rodríguez y Alvarado-Ortega, 2021b |
Fm. Cerro del Pueblo (Coahuila), México |
Campaniano tardío |
|
Tsintaosaurus spinorhinus Young, 1958 |
Fm. Jingangkou (Shandong), China |
Campaniano temprano |
|
Velafrons coahuilensis Gates, Sampson, Delgado de Jesús, Zanno, Eberth, Hernández-Rivera, Aguillón-Martínez y Kirkland, 2007 |
Fm. Cerro del Pueblo (Coahuila), México |
Campaniano tardío |
|
Wulagasaurus dongi Godefroit, Hai, Yu y Lauters, 2008 |
Fm. Yuliangze (Heilongjang), China |
¿Maastrichtiano tardío? |
|
Xuwulong yueluni You, Li y Liu, 2011 |
Gpo. Xinminpu (Gansu), China |
Aptiano tardío |
|
Yamatosaurus izanagii Kobayashi, Takasaki, Kubota y Fiorillo, 2021 |
Fm. Kita-ama (Isla Awaji), Japón |
Maastrichtiano temprano |
|
Yunganglong datongensis Wang R, You, Xu, Wang S, Yi, Xie, Jia y Li, 2013 |
Fm. Zhumapu (Shanxi), China |
Cenomaniano |
|
Zhanghenglong yangchengensis Xing, Wang, Han, Sullivan, Ma, He, Hone, Yan, Du y Xu, 2014b |
Fm. Majiacun (Henan), China |
Santoniano medio |
|
Zuoyunlong huangi Wang R, You, Wang S, Xu, Yi, Xie, Jia y Xing, 2015 |
Fm. Zhumapu (Shanxi), China |
Cenomaniano |
Tabla 1 (continuación). Lista de Hadrosauroidea válidos
Tabla 1 (continuación). Lista de Hadrosauroidea válidos
Tabla 2. Glosario de términos y definiciones usados para la descripción de estructuras osteológicas.
|
Término en castellano (inglés) |
Definiciones |
|
TÉRMINOS DE PARTES DEL HUESO |
|
|
Ángulo (angle) |
Esquina de una estructura anatómica, punto de unión de dos bordes o superficies. |
|
Ápice (apex) |
La parte superior de un cuerpo o la punta de un extremo cónico. |
|
Área o región (area, region) |
Región o zona de una superficie de un órgano con una función específica. |
|
Base (base, basis) |
Estructura anatómica que funciona como soporte fundamental. |
|
Borde (border, margin) |
Extremo de cualquier órgano anatómico, articular o no. |
|
Cara (facie) |
Cualquier superficie o cara de un hueso. |
|
Cara articular (articular facie) |
Superficie articular de los huesos o cartílagos que forman una unión con otro. |
|
Cuerpo (centrum, body, central part) |
La parte principal de un órgano o región anatómica. |
|
Expansión (expansion) |
Una región, superficie o área de mayor volumen. |
|
Plano (plane) |
Superficie plana de un hueso. |
|
Plataforma (platform) |
Superficie horizontal plana, más alta que el nivel de las áreas a su alrededor. |
|
TÉRMINOS DE FORMA |
|
|
Ala (ala, wing) |
Cualquier parte anatómica en forma de ala. |
|
Arco (arch) |
Cualquier estructura anatómica con forma de arco. |
|
Escama (squama) |
La parte plana de un hueso, especialmente las partes de la cavidad craneal. |
|
Lámina (lamina, plate) |
Término genérico que denota varias cosas planas. |
|
Labio (labrum, lip) |
Término usado para denotar cualquier estructura anatómica parecida o semejante a un labio. |
|
Quilla o carina (quill, carina) |
Eminencia de órgano con forma de quilla de un bote. |
|
Septo o tabique (septum) |
Término genérico de lámina que divide una cavidad en dos partes. |
|
PROYECCIONES ARTICULARES |
|
|
Cabeza (caput, head, capitulum) |
Eminencia redondeada o cualquier parte anatómica que parezca cabeza. |
|
Cigoapófisis (zygapophysis) |
Articulaciones de las vértebras. |
|
Cóndilo (condyle) |
Eminencia articular redondeada o elipsoidal que tiene una superficie convexa formando una articulación tipo condilar. |
|
Hipósfeno (hyposphene) |
Proyección triangular pequeña, de la base caudal del proceso espinoso (Stefanic y Nesbitt, 2019) |
|
Pedúnculo (peduncle) |
Cualquier parte angosta, con tamaños y longitudes variables, por la cual se unen dos huesos o regiones de éstos. |
|
Tróclea (trochlea) |
Proyección articular lisa con un surco en el centro, delimitada por dos paredes. |
|
Yugo (jugum, yoke ) |
Cualquier cresta o surco que conecta a otra estructura anatómica. |
|
PROYECCIONES NO ARTICULARES |
|
|
Cuello (neck) |
Estructura angosta o constreñida que se encuentra entre dos articulaciones o dos estructuras anatómicas. |
|
Cresta (crista, crest, ridge) |
Elevación estrecha y elongada. Particular en huesos donde se unen tendones y músculos. |
|
Eminencia (eminence) |
Cualquier proyección del hueso no tan prominente como el proceso. |
|
Epicóndilo (epicondyle) |
Proyección adyacente al cóndilo. |
|
Escapo (scapus, shaft, stem) |
Estructura delgada y atenuada del hueso, en lugar del cuerpo anatómico, cuando no es posible usarlo (Baumel y Witmer, 1993). |
|
Espina (spine) |
Proyección aguda, filosa, larga y plana. |
|
Línea (line, ridge) |
Elevación linear baja y angosta, no tan afilada como la cresta. |
|
Maléolo (malleolus) |
Cada una de las eminencias óseas redondeadas, internas y externas, del extremo inferior de la pierna o tobillo. |
|
Proceso o apófisis (process) |
Cualquier proyección que sale de la sección principal en un hueso. |
|
Rama (ramus) |
Cualquier subdivisión, sección o bifurcación. Hendrickx y Mateus (2014) recomiendan usarlo en lugar de “proceso” cuando son estructuras grandes, complejas y que contienen multiples procesos. |
|
Torus o intumescencia (torus, swelling) |
Eminencia redondeada y lisa de un cuerpo, o cualquier cresta pequeña ósea que se observa en el cráneo. |
|
Tubérculo (tubercle, tuberculum) |
Pequeña proyección, elevación redondeada, lisa o rugosa de un hueso. |
|
Tuberosidad (tuberosity, tuber) |
Proyección larga, ovoide y de superficie rugosa. Sirve de unión de músculos y ligamentos. |
|
Trocánter (trochanter) |
Grandes procesos rugosos y planos, únicos del fémur. |
|
DEPRESIONES ARTICULARES |
|
|
Cótilo (cotyla) |
Superficie articular cóncava y poco profunda (Baumel y Witmer, 1993). |
|
Cavidad glenoidea (glenoid cavity) |
Fosa poco profunda de la escápula. |
|
Acetábulo (acetabulum) |
Fosa profunda de la cintura pélvica, con forma de copa, que recibe la cabeza femoral. |
|
DEPRESIONES NO ARTICULARES |
|
|
Concavidad (concavity) |
Área deprimida, más pequeña que una fosa, con limites definidos, de una superficie de un órgano o estructura. |
|
Depresión (depression) |
Área deprimida, desplazamiento hacia abajo o hacia dentro de un órgano o estructura. Puede ser más grande o no que una fosa y tiene límites difusos, a diferencia de la fosa. |
|
Escotadura o incisura (cleft, notch) |
Muesca que puede ser articular o no. |
|
Excavación (excavation) |
Proceso de formación de una caverna en huesos. |
|
Fosa (fossa) |
Depresión ovalada de cualquier cuerpo anatómico, usualmente, es larga, amplia y poco profunda. |
|
Fóvea (fovea) |
Depresión pequeña circular, normalmente, en la superficie lateral de un hueso largo. |
|
Impresión (impression) |
Marca, excavación o depresión en el cual se fija un ligamento o músculo. |
|
Receso (recess) |
Depresión, fosa o cavidad sin salida donde hay una formación anatómica importante, como un órgano blando. |
|
Surco (sulcus, groove) |
Depresión larga y estrecha. |
|
HUECOS |
|
|
Abertura (aperture) |
Agujero que funcione como salida o comunicación de un ducto o cavidad amplia y de gran tamaño. |
|
Alveólo (alveolus) |
Cuenca donde se inserta un diente. |
|
Canal (canal) |
Cualquier formación anatómica en forma de tubo que tiene sus propias paredes de estructura compacta y sirve de paso de fluidos u otros órganos. |
|
Cavidad (cavity) |
Cualquier espacio cerrado en el cuerpo. |
|
Cámara (chamber) |
Cavidad relativamente pequeña. |
|
Foramen (foramen, foramina) |
Cualquier orificio pequeño o hueco por donde pasan músculos, arterias, venas, nervios u otras estructuras. |
|
Foramen nutricio (nutriet foramen) |
Agujero de los huesos por donde pasan los vasos sanguíneos. |
|
Foramen pneumático (pneumatic foramen) |
Orificio que permite el paso del aire al interior de un hueso (Currie y Padian, 1997). |
|
Fenestra (fenestra) |
Ventana. Cualquier orificio de un hueso o entre huesos que está cerrado por una membrana. |
|
Fontanela o fontículo (fontanelle) |
Nombre genérico de una zona membranosa interpuesta entre huesos del cráneo. |
|
Seno (sinus) |
Cámara, canal o espacio dentro del hueso normalmente ocupado por aire. |
Términos de Baumel y Witmer, 1993. Definiciones de Dorian (1988), Frandson et al. (2009), Dorland (2012) y White et al. (2012).
Tabla 2 (continuación). Glosario de términos y definiciones usados para la descripción de estructuras osteológicas.
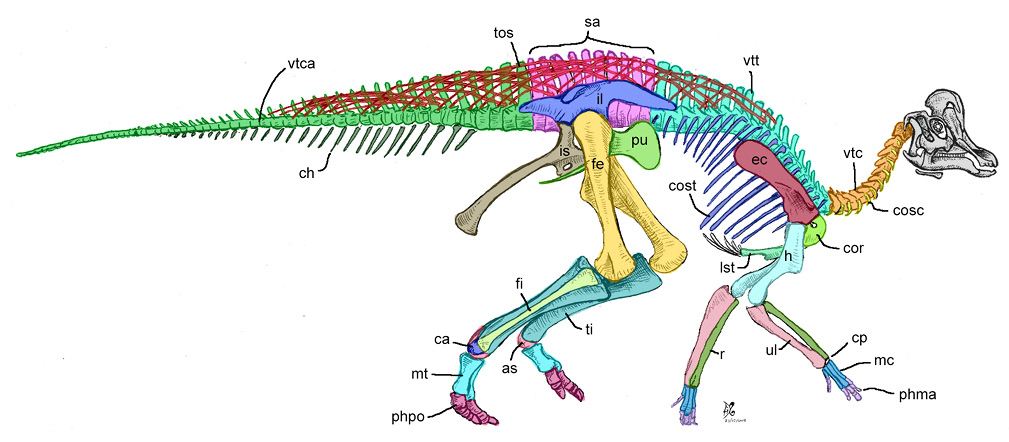
Tabla 2 (continuación) – Glosario de términos y definiciones usados para la descripción de estructuras osteológicas.
Figura 2. Reconstrucción del esqueleto de Velafrons coahuilensis (CPC-59), se muestran los distintos huesos que conforman el esqueleto postcraneano de los hadrosauroideos.
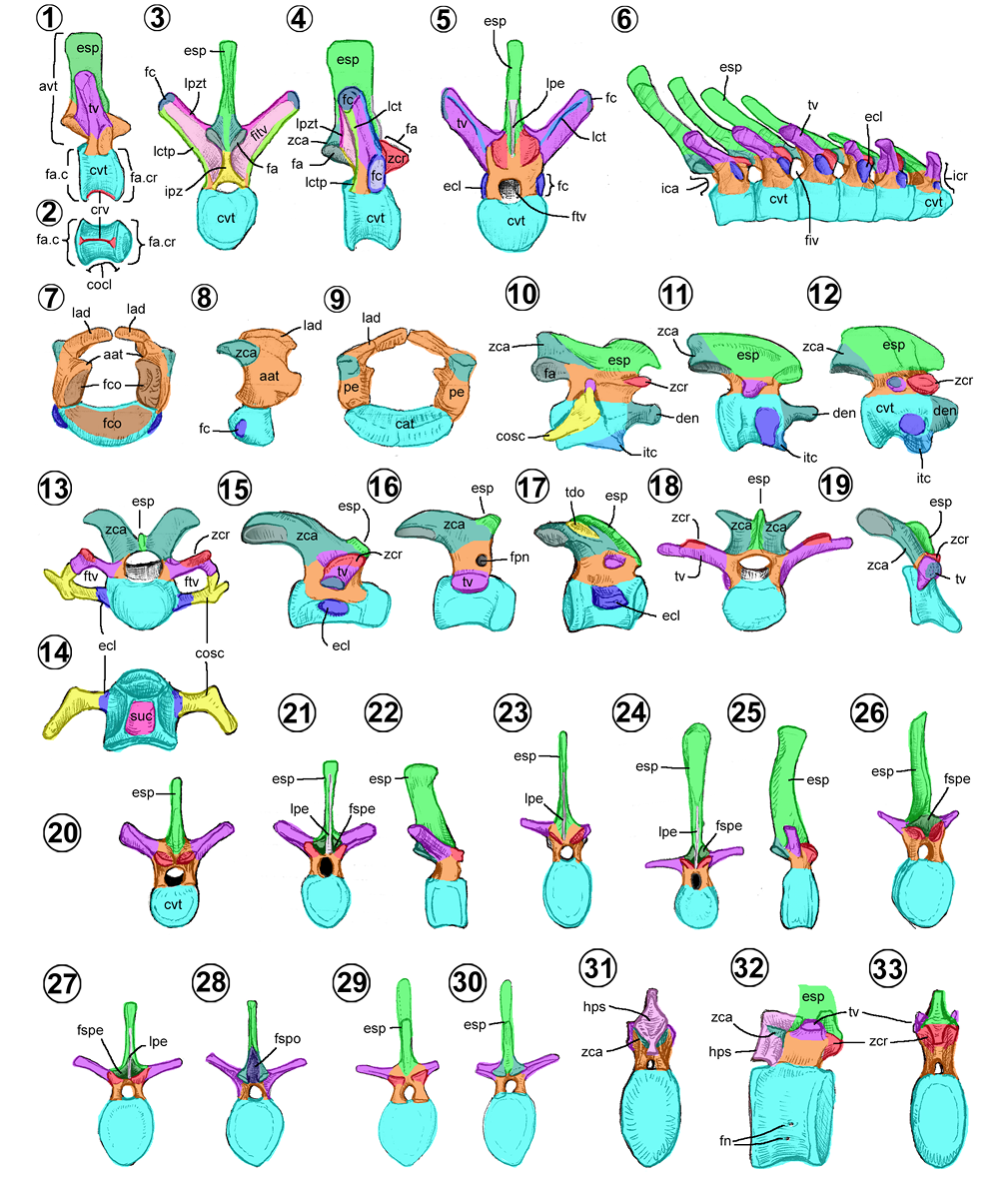
Figura 3. Vértebras cervicales y torácicas de hadrosauroideos. 1–5, Vértebra torácica anterior de Edmontosaurus (Campione, 2014, figs. 13.4K–M, O). 6, Serie de vértebras torácicas anteriores articuladas de Magnapaulia (IGM 5844). 7–9, Axis de Plesiohadros (Tsogtbaatar et al., 2014, figs. 7.19A–B, E). Atlas de: 10, Brachylophosaurus (Prieto-Márquez, 2001, fig. 42), 11, Huehuecanauhtlus (IGM 6253) y 12, Hadrosauroideo de Woodbine (Main et al., 2014, fig. 5.4B). Vértebra cervical media de: 13–14, Edmontosaurus (Campione, 2014, figs. 13.2F, O) y 15, Brachylophosaurus (Prieto-Márquez, 2001, fig. 45). 16, Cervical de Lapampasaurus (Coria et al., 2012, fig. 2.5). 17, Cervical de Equijubus (McDonald et al., 2014, fig. 3.11C). 18–19, Cervical posterior de Edmontosaurus (Campione, 2014, figs. 13.2K, M). Torácica media de: 20, Eolambia (McDonald et al., 2012, fig. 23H), 21–22, Bactrosaurus (Godefroit et al., 1998, pls. 8.3A–B) y 23, Eotrachodon (Prieto-Márquez et al., 2016, fig. 23E). Torácica posterior de: 24–25, Barsboldia (Maryanska y Osmólska, 1981, pls. 14.2b–2c) y 26, Gryposaurus (USNM V8058). 27–28, Torácica media de Edmontosaurus (Campione, 2014, figs. 13.6F–H). 29–30, Torácica posterior de Gryposaurus (Freedman, 2015, figs. 5.2E–F). 31–33, Torácica media de Equijubus (McDonald et al., 2014, figs. 3.14A–C). Dibujos sin escala en vistas lateral (1, 4, 6, 8, 10–12, 15-17, 19, 22, 25, 32), ventral (2, 14), caudal (3, 9, 28, 30, 31) y craneal (5, 7, 13, 18, 20, 21, 23, 24, 26, 27, 29, 33).
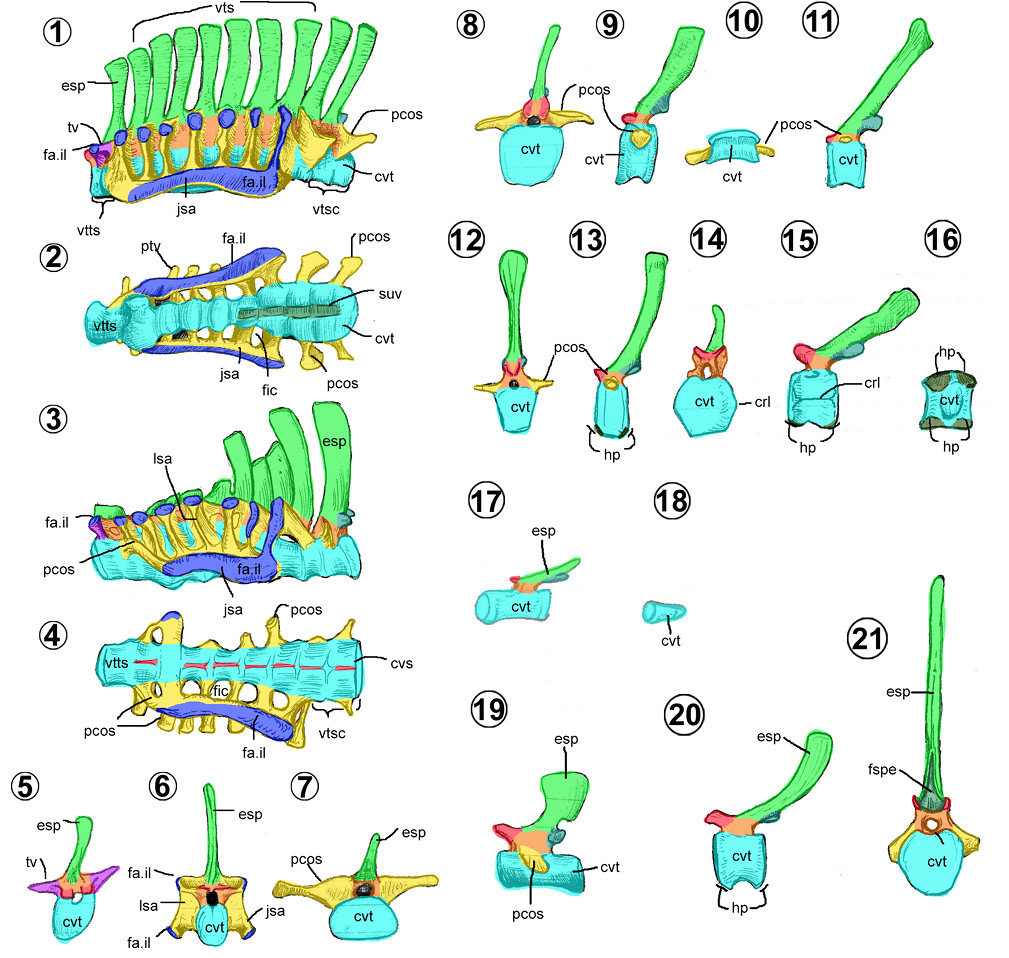
Figura 4. Sacro y vértebras caudales de hadrosauroideos. Sacro de: 1–2, Shantungosaurus (Hu et al., 2001, fig. 2.16) y 3–4, Tsintaosaurus (Young, 1958, fig. 17). 5, Vértebra toracosacral de Edmontosaurus (Campione, 2014, fig. 13.9A). 6, Vértebra sacra de Magnapaulia (Prieto-Márquez et al., 2012, fig. 10C). 7, Caudosacral de Edmontosaurus (Campione, 2014, fig. 13.9D). 8–10, Caudales anteriores de Edmontosaurus (Campione, 2014, figs. 13.10F–G, J). 11, Caudal media de: Olorotitan (Godefroit et al., 2012b, figs. 17B–C) y 12–13, Barsboldia (Maryanska y Osmólska, 1981, pl. 18). Caudales posteriores de: 14–16, Edmontosaurus (Campione, 2014, figs. 13.11F–G, J) y 17–18, Hadrosáurido de Dinosaur Park (Allen, 2009, fig. 91). 19, Caudal anterior de Tethyshadros (Dalla Vecchia, 2009, fig. 4). 20, Caudal posterior de Arenysaurus (Dalla Vecchia, 2020, fig. 13). 21, Caudal anterior de Magnapaulia (Prieto-Márquez et al., 2012, fig. 11C). Dibujos sin escala en vista lateral (1, 3, 9, 11, 13, 15, 17–20), ventral (2, 4, 10, 16), craneal (5, 6, 8, 12, 14, 21) y caudal (7).
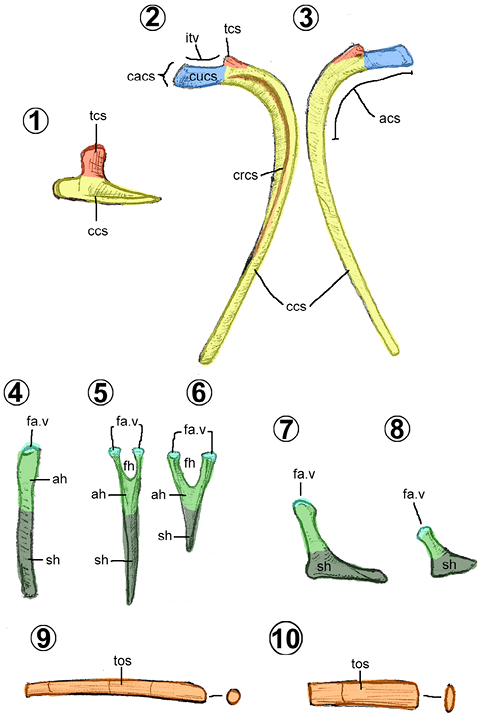
Figura 5. Costillas, cheurónes y tendones osificados de hadrosauroideos. 1, Costilla cervical de Kamuysaurus (Kobayashi et al., 2019, fig. S15h). 2-3, Costilla torácica de Eolambia (McDonald et al., 2012, fig. 24). Cheurón de: 4–6, Eolambia (McDonald et al., 2012, figs. 26H–J) y 7–8, Tethyshadros (Dalla Vecchia, 2009, fig. 4B). 9–10, Tendones osificados de un hadrosáurido de Dinosaur Park (Allen, 2009, fig. 112). Dibujos sin escala en vista lateral (1, 4, 7, 8, 9, 10), craneal (2) y caudal (3, 9, 10).
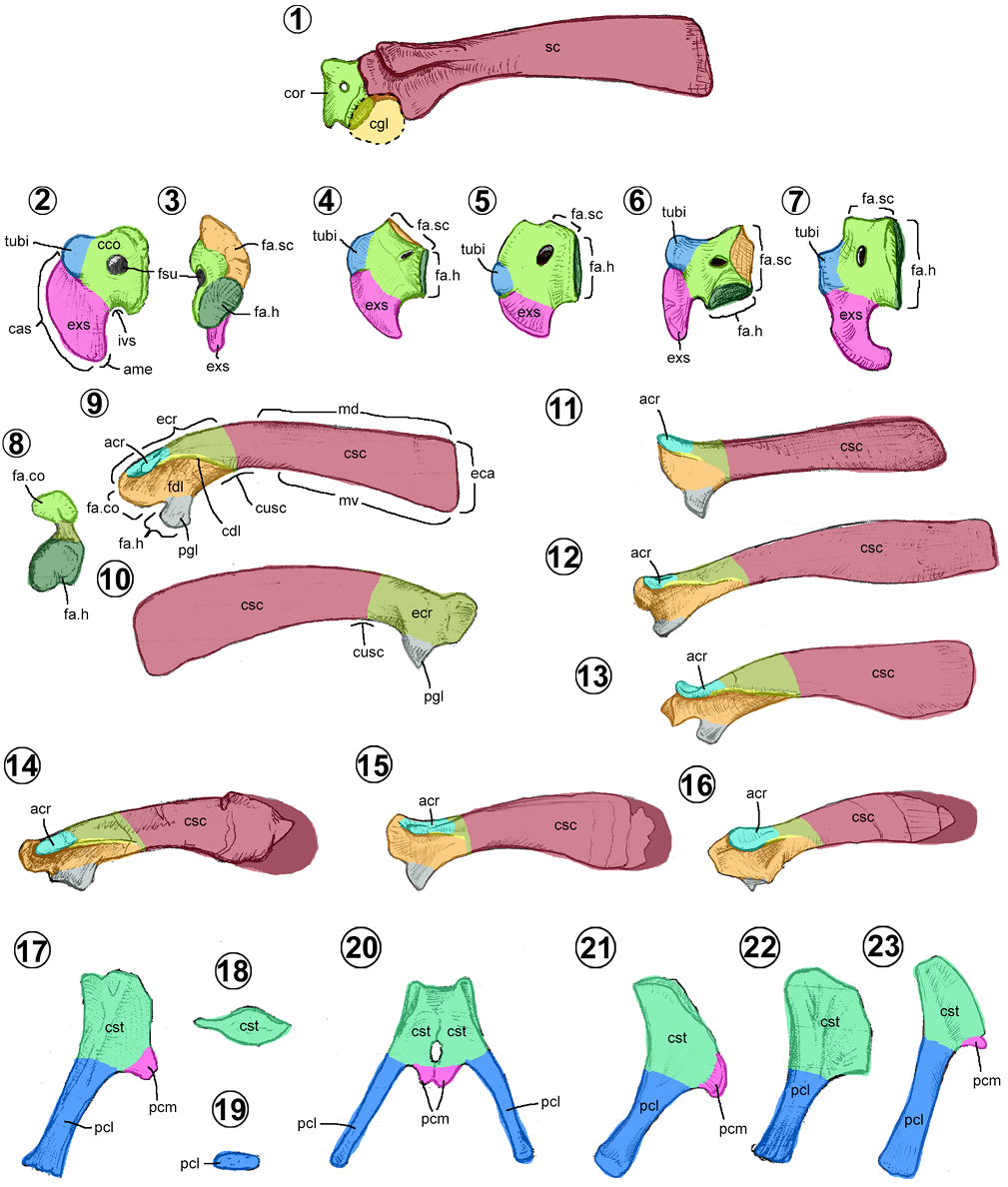
Figura 6. Cintura pectoral en hadrosauroideos. 1, Escápula y coracoides articulados de Gobihadros (Tsogtbaatar et al., 2019, fig. 22A). Coracoides de: 2–3, Edmontosaurus (Campione, 2014, figs. 13.13F, H), 4, Jinzhousaurus (Wang et al., 2011, fig. 6b), 5, Gilmoreosaurus (Prieto-Márquez y Norell, 2010, fig. 11A), 6, Amurosaurus (Godefroit et al., 2004, fig. 13A) y 7, Brachylophosaurus (Prieto-Márquez, 2008, fig. H.6C). Escápulas de: 8–10, Edmontosaurus (Campione, 2014, figs. 13.13A–C), 11, Eolambia (McDonald et al., 2012, fig. 27C), 12, Corythosaurus (Prieto-Márquez, 2008, fig. H.9B), 13, Parasaurolophus (Prieto-Márquez et al., 2019a, fig. 12D), 14, hadrosaurio de Sabinas (PASAC-1), 15, Hypacrosaurus (Prieto-Márquez et al., 2019a, fig. 12E) y 16, Adynomosaurus (Prieto-Márquez et al., 2019a, fig. 6A). Placas esternales de: 17–20, Edmontosaurus (Campione, 2014, figs. 13.13I–J, L; Prieto-Márquez, 2014, fig. 7C), 21, Equijubus (McDonald et al., 2014, fig. 3.17B), 22, Batyrosaurus (Godefroit et al., 2012c, fig. 20.11A) y 23, Brachylophosaurus (Prieto-Márquez, 2001, fig. 59). Dibujos sin escala en vista lateral (1–2, 4–7, 9, 11–16), caudal (3, 19), medial (10), craneal (8, 18) y ventral (17, 20–23).
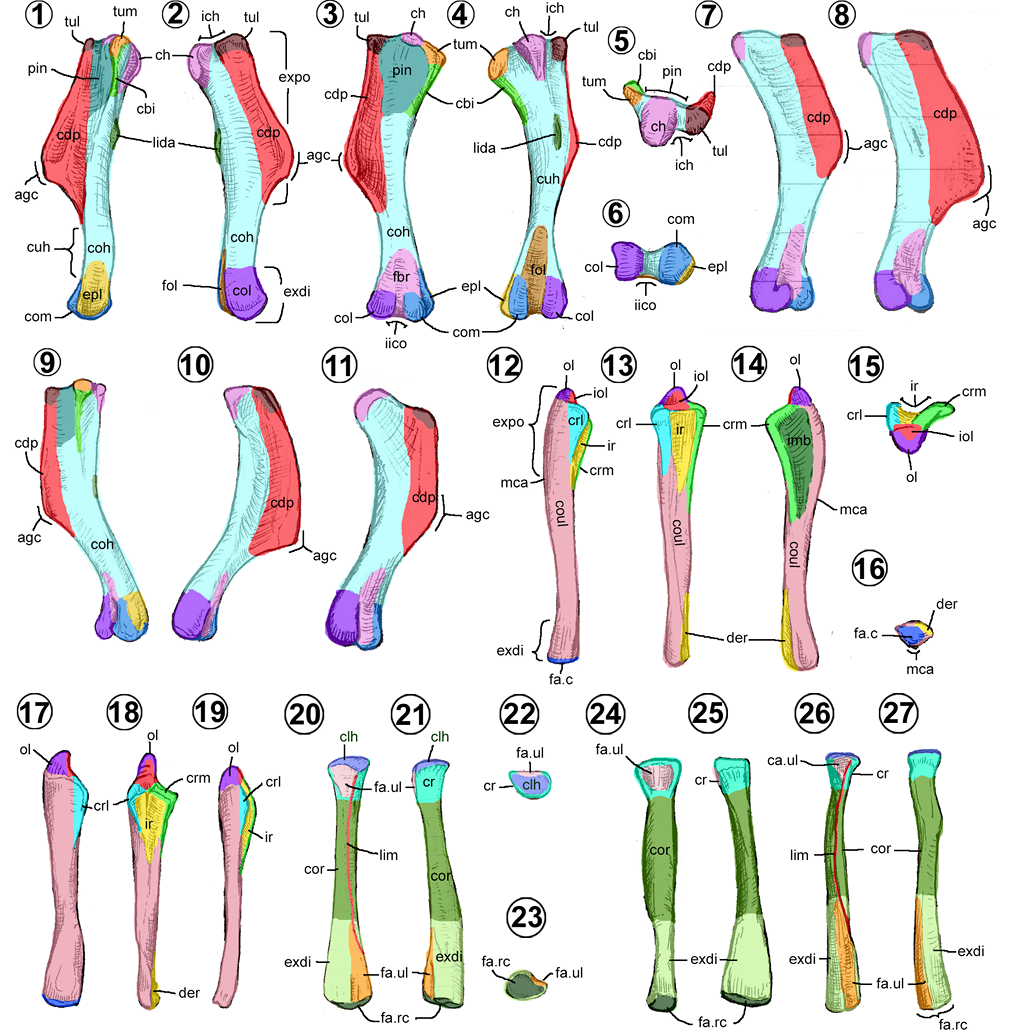
Figura 7. Brazo y antebrazo de hadrosauroideos. Húmeros de: 1–6, restaurado Edmontosaurus (Campione, 2014, figs. 13.14A–B, G, H, K), 7, Hadrosaurus (Prieto-Márquez, 2011, fig. 3), 8, Magnapaulia (Prieto-Márquez et al., 2012, fig. 14G), 9, Gryposaurus (Prieto-Márquez, 2012, fig. 11B), 10, Olorotitan (Godefroit et al., 2012a, fig. 19A1), y 11, Gilmoreosaurus (Prieto-Márquez y Norell, 2010, fig. 12B). Ulnas de: 12–16, Gryposaurus (Freedman, 2015, fig. 5.10), 17–18, Eolambia (McDonald et al., 2012, figs. 28E, G) y 19, Lambeosaurino de Els Nerets (Conti et al., 2020, fig. 6D). Radios de: 20–23, hadrosaurio HB (sin número), 24–25, Eolambia (McDonald et al., 2012, figs. 28L–M), 26, Kerberosaurus (Godefroit et al., 2012b, fig. 17C) y 27, Edmontosaurus (Campione, 2014, fig. 13.16C). Dibujos sin escala en vista medial (1, 9, 14), lateral (2, 7–8, 10–11, 12, 17, 19, 21, 25, 27), craneal (3, 13, 18), caudal (4, 20, 24, 26), proximal (5, 15, 22) y distal (6, 16, 23).
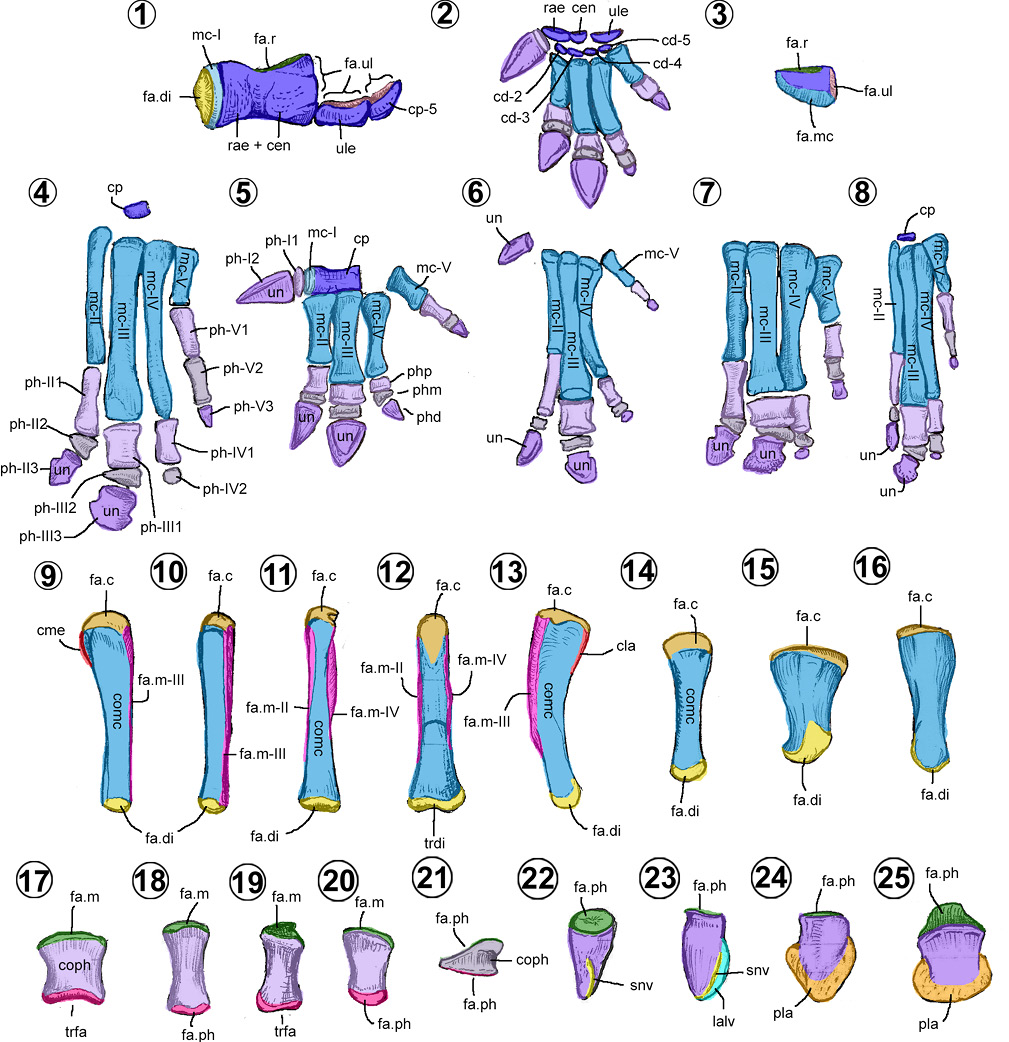
Figura 8. Mano de hadrosauroideos. 1, Carpo reconstruido de Jinzhousaurus (Wang et al., 2011, fig. 8). 2, Mano reconstruida de Bolong (Wu y Godefroit, 2012, fig. 19.9; Zheng et al., 2014, fig. 12). 3, Carpiano de Edmontosaurus (Prieto-Márquez, 2014, fig. 10C). Mano de: 4, Edmontosaurus (Prieto-Márquez, 2014, fig. 10A), 5, Jinzhousaurus (Wang et al., 2011, fig. 8), 6, Probactrosaurus (Norman, 2002, fig. 26A), 7, Parasaurolophus (Parks, 1922, pl. IX-1) y 8, Brachylophosaurus (Hohloch, 2013, fig. 18). Metacarpiano-II de: 9, Eolambia (McDonald et al., 2012, fig. 29A) y 10, Probactrosaurus (Norman, 2002, fig. 24A). Metacarpiano-III de: 11, Probactrosaurus (Norman, 2002, fig. 24C) y 12, Eolambia (McDonald et al., 2012, fig. 29G). 13, Metacarpiano-IV de Eolambia (McDonald et al., 2012, fig. 29N). Metacarpiano-V de: 14, Eolambia (McDonald et al., 2012, fig. 29P), 15, Bactrosaurus (Prieto-Márquez, 2008, fig. H.24A) y 16, Maiasaura (Prieto-Márquez, 2008, fig. H.24B). 17, Falange manual III-1 y 18, Falange IV-1 de Eolambia (McDonald et al., 2012, figs. 30B, F). 19, Falange IV-1 de Kamuysaurus (Kobayashi et al., 2019, fig. S19g). 20, Falange V-1 de Eolambia (McDonald et al., 2012, fig. 30M). 21, Falange III-2 de un hadrosáurido del Dinosaur Park (Allen, 2009, fig. 102A1). 22, Ungueal manual-1 o póllex de Eolambia (McDonald et al., 2012, fig. 30A). 23, Ungueal-II de Probactrosaurus (Norman, 2002, fig. 25H). 24, Ungueal-II y 25, ungueal-III de Brachylophosaurus (Prieto-Márquez, 2001, fig. 71). Dibujos sin escala en vista dorsal.
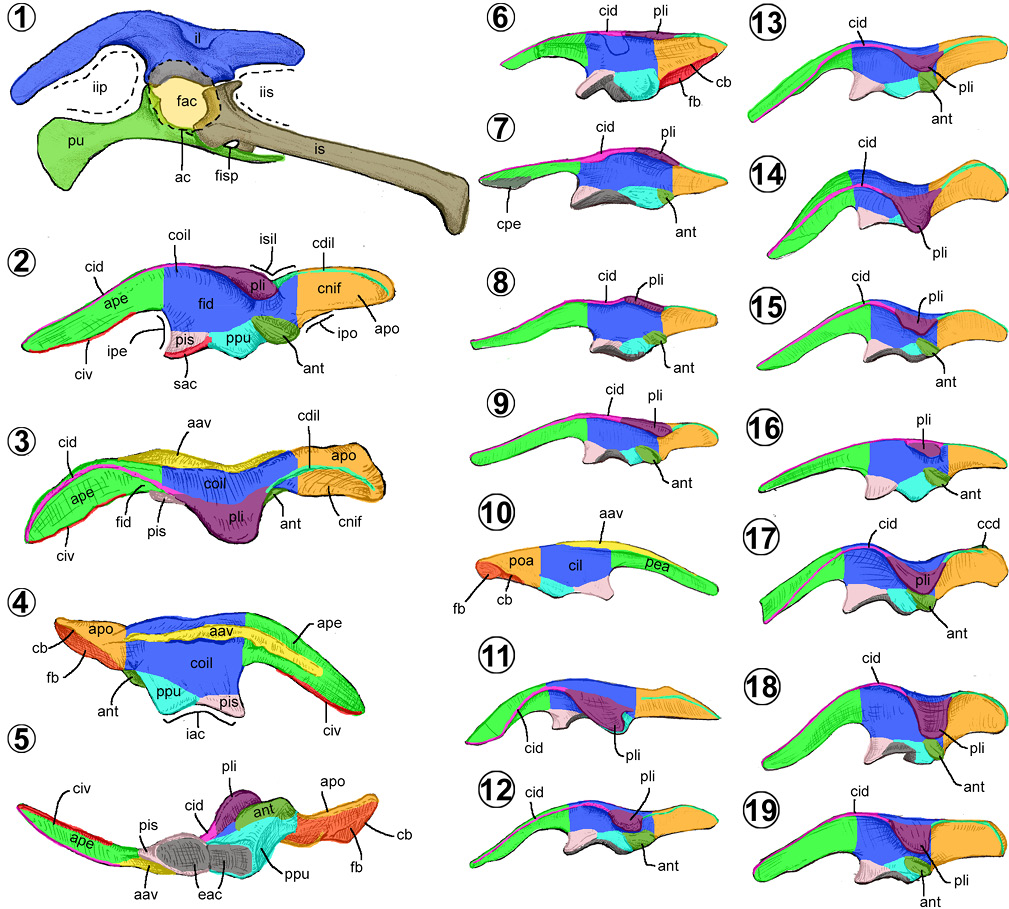
Figura 9. Ilion de hadrosauroideos. 1, Cintura pélvica de Corythosaurus (Brown, 1916, figs. 2, 4; Horner et al., 2004, fig. 20.12B). Ilion de: 2–5, Velafrons (CPC-59), 6, Jinzhousaurus (Wang et al., 2011, fig. 9a), 7, Eolambia (McDonald et al., 2017, fig. 9A), 8, Tanius (Borinder et al., 2021, fig. 9C), 9–10, Gilmoreosaurus (Prieto-Márquez y Norell, 2010, figs. 13D–E), 11, Tethyshadros (Dalla Vecchia, 2009, fig. 5B), 12, Edmontosaurus (Prieto-Márquez, 2014, fig. 11D), 13, kritosaurino de Careless Creek (Prieto-Márquez y Gutarra, 2016, fig. 6.1), 14, Prosaurolophus (Drysdale et al., 2019, fig. 9K), 15, Brachylophosaurus (Prieto-Márquez, 2001, fig. 60), 16, Sahaliyania (Godefroit et al., 2008, fig. 10A1), 17, Hypacrosaurus (USNM V11950), 18, Parasaurolophus (Ostrom, 1963, fig. 93b) y 19, Tsintaosaurus (Young, 1958, fig. 26.2). Dibujos sin escala en vista lateral (1–2, 6-9, 11–19), dorsal (3), medial (4, 10), y ventral (5).
Tabla 3. Morfotipos iliacos de hadrosauroideos.
|
Morfotipos |
Cuerpo ilíaco |
Proceso lateral |
Antitrocánter |
Ala preacetabular |
Ejemplo |
|
1 |
Más alto que largo, con borde dorsal recto a ligeramente convexo |
Largo y sinuoso |
Ausente |
Triangular |
Equijubus |
|
2 |
Más largo que alto |
Largo y sinuoso a cuneiforme |
Incipiente |
Triangular a oblonga |
Gilmoreosaurus |
|
3 |
Más largo que alto, comparativamente más bajo |
Acampanado y linguliforme |
Presente y desarrollada |
Oblonga |
Edmontosaurus |
|
4 |
Más alto que largo |
Oblonga e inclinada dorsalmente |
Gryposaurus |
||
|
5 |
Trapeziforme |
Brachylophosaurus |
|||
|
6 |
Más alto que largo, comparativamente más largo |
Acampanado y relativamente pequeño |
Oblonga |
Corythosaurus |
|
|
7 |
Más alto que largo, comparativamente más robusto |
Linguliforme y alto |
Oblonga y profunda |
Parasaurolophus |
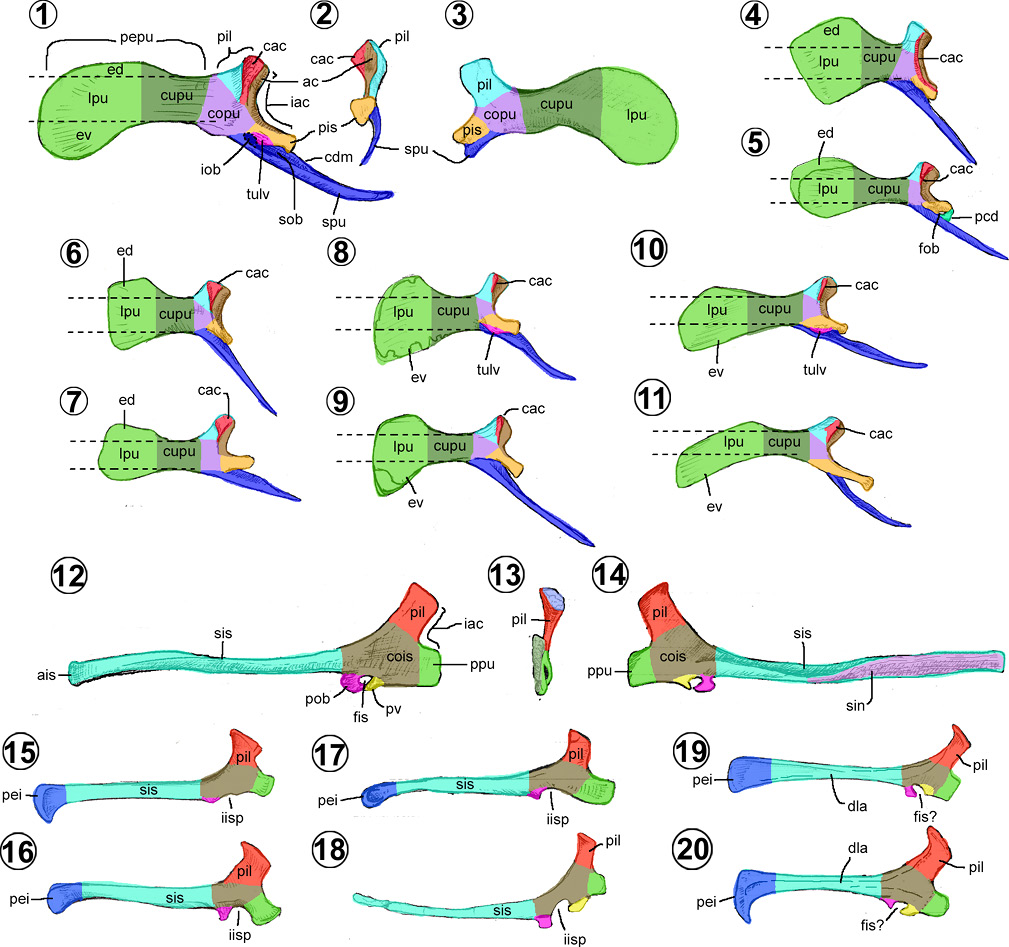
Figura 10. Pubis e isquion de hadrosauroideos. Pubis de: 1–3, Edmontosaurus (Campione, 2014, fig. 13.18A–C), 4, Gobihadros (Tsogtbaatar et al., 2019, fig. 28A), 5, Eolambia (McDonald et al., 2017, fig. 9E), 6, Parasaurolophus (Ostrom, 1963, fig.), 7, Sahaliyania (Godefroit et al., 2008, fig. 10C1), 8, Hypacrosaurus (Guenther, 2009, fig. 10C), 9, Saurolophus (Brown, 1913, fig. 1), 10, Brachylophosaurus (Prieto-Márquez, 2001, fig. 61) y 11, Secernosaurus (Prieto-Márquez y Salinas, 2010, fig. 16A). Isquiones de: 12–14, Edmontosaurus (Campione, 2014, fig. 13.19A–B), 15, Eolambia (McDonald et al., 2012, fig. 31F), 16, Gilmoreosaurus (Prieto-Márquez y Norell, 2010, fig. 14A), 17, Hadrosauroideo de Woodbine (Main et al., 2014, fig. 5.11C), 18, Hadrosaurio de Sabinas (PASAC-1), 19, Corythosaurus (USNM PAL358560) y 20, Parasaurolophus (Ostrom, 1963, fig. 90a). Dibujos sin escala en vista lateral (1, 4–12, 15–20), caudal (2), medial (3, 14) y craneal (13).
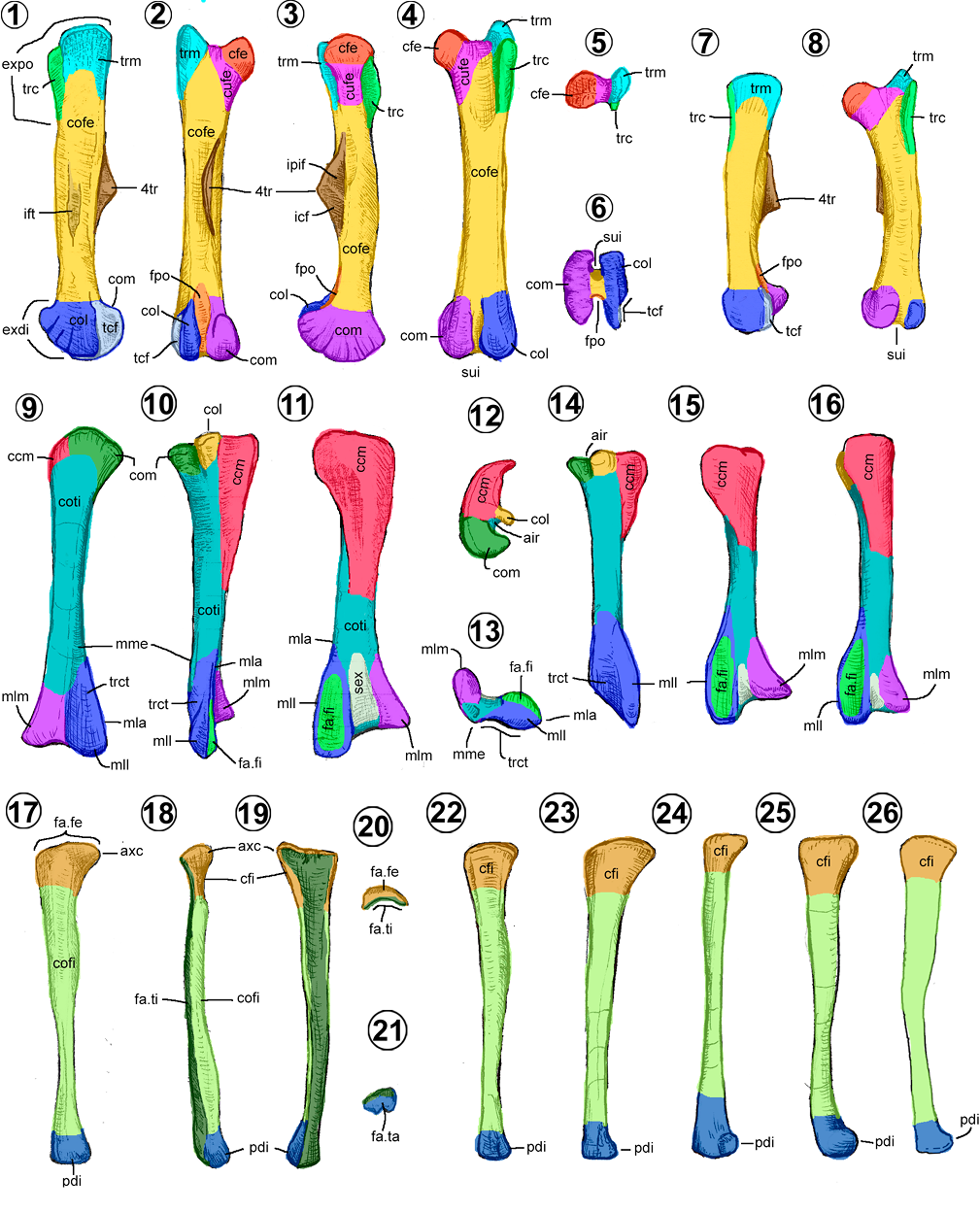
Figura 11. Miembro pélvico de hadrosauroideos. Fémures de: 1, 3, 5–6, Edmontosaurus (Campione, 2014, fig. 13.20A–B, C, F), 2, 4, Kamuysaurus (Kobayashi et al., 2019, fig. S21a, c) y 7–8, Choyrodon (Gates et al., 2018, fig. 19D). Tibias de: 9–13, Kamuysaurus (Kobayashi et al., 2019, fig. S22a–f), 14–15, Gilmoreosaurus (Prieto-Márquez y Norell, 2010, fig. 16A–B) y 16, Amurosaurus (Godefroit et al., 2004, fig. 16A2). Fíbulas de: 17–21, Edmontosaurus (Campione, 2014, fig. 13.22A–E), 22, Eolambia (McDonald et al., 2012, fig. 32M), 23, Brachylophosaurus (Prieto-Márquez, 2001, fig. 75), 24, Gryposaurus (Prieto-Márquez, 2012, fig. 13A), 25, Amurosaurus (Godefroit et al., 2004, fig. 16B2) y 26, Parasaurolophus (Ostrom, 1963, fig. 92d). Dibujos sin escala en vista lateral (1, 7, 10, 14, 17, 22–26), caudal (2, 9), medial (3, 19), cranial (4, 8, 11, 15–16, 18), proximal (5, 12, 20) y distal (6, 13, 21).
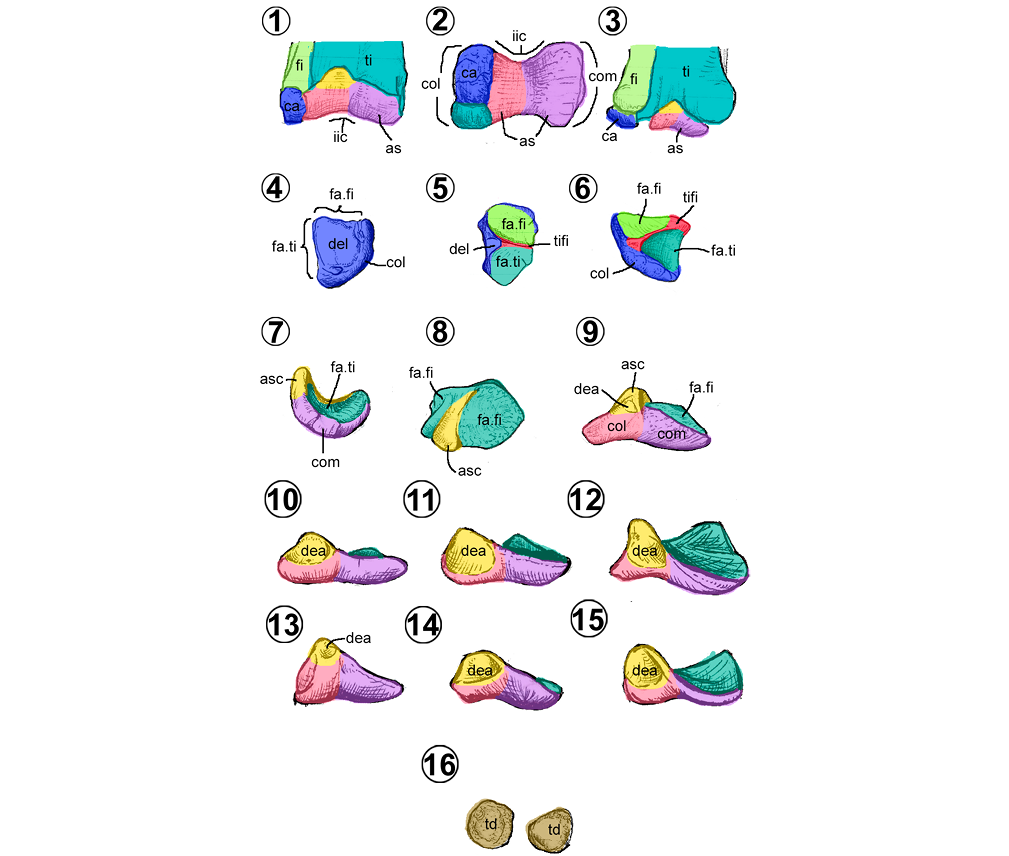
Figura 12. Tarso de hadrosauroideos: tarsales proximales articulados con la tibia y fíbula de: 1–2, Brachylophosaurus (Prieto-Márquez, 2001, figs. 77–78) y 3, Tsintaosaurus (Prieto-Márquez, 2008, fig. J.5F). 4–6, Calcáneo del hadrosaurio HB (sin número). Astrágalo de: 7–8, Saurolofino de McCone (Zheng et al., 2011 fig. 7A, H), 9, Edmontosaurus (Campione, 2014, fig. 13.22I), 10, Gryposaurus (Freedman, 2015, fig. 5.20B), 11, Probrachylophosaurus (Freedman y Horner, 2015, fig. 21F), 12, Bonapartesaurus (Cruzado-Caballero y Powell, 2017, fig. 11B), 13, Parasaurolophus (Prieto-Márquez, 2008, fig. J.5D), 14, Amurosaurus (Godefroit et al., 2004, fig. 17A2) y 15, Charonosaurus (Godefroit et al., 2001, fig. 22A). 16, Tarsal distal de Brachylophosaurus (Prieto-Márquez, 2001, fig. 79). Dibujos sin escala en vista craneal (1, 3, 9–15), distal (2), lateral (4), proximal (5, 8, 16) y medial (6, 7).
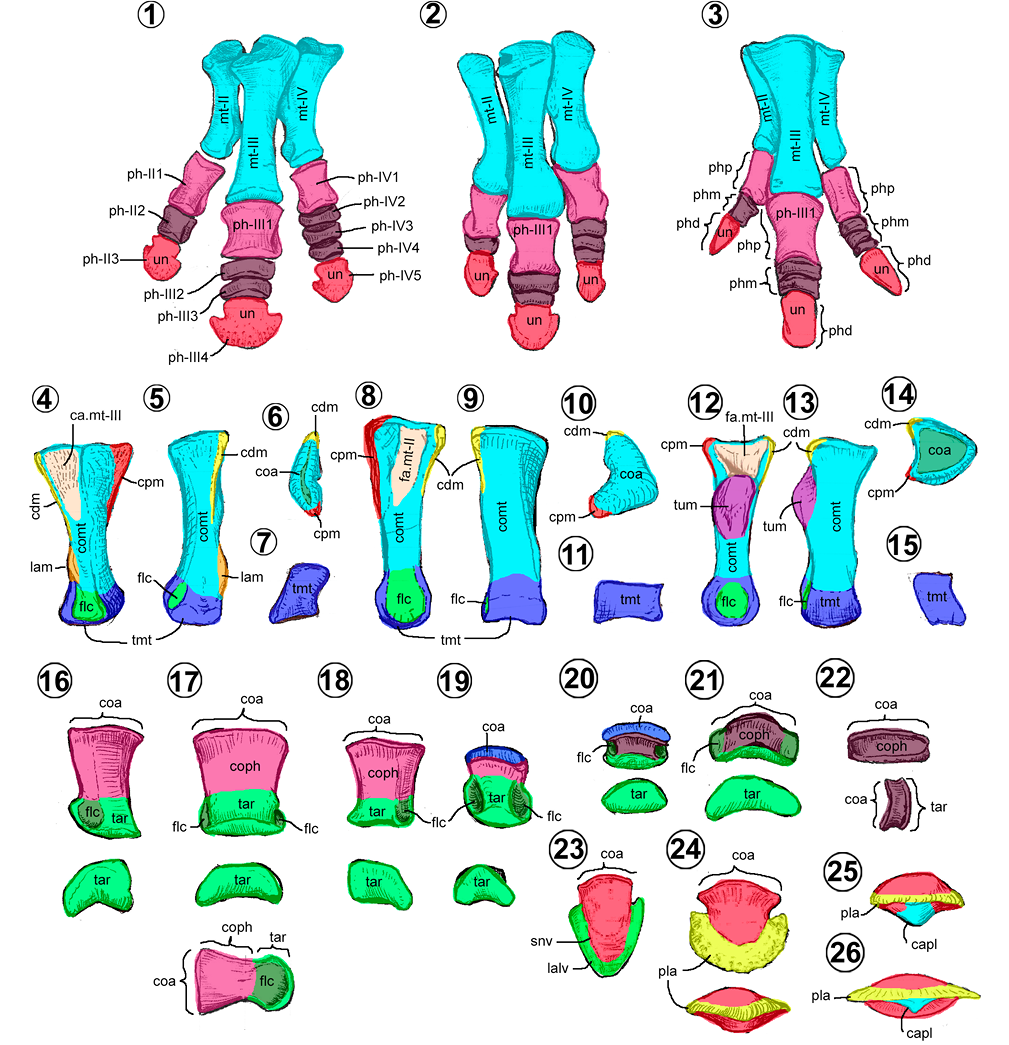
Figura 13. Pie de hadrosauroideos. Pie de: 1, Edmontosaurus (Prieto-Márquez, 2014, fig. 14B), 2, Nipponosaurus (Nagao, 1983, pl. 17) y 3, Probactrosaurus (Norman, 2002, fig. 33). 4–7, Metatarsiano-II, 8–11, metatarsiano-III y 12–15, metatarsiano-IV del hadrosaurio HB (sin número). 16, Falange pedal II-1, 17, falange III-1 y 18, falange IV-1 de Edmontosaurus (Zheng et al., 2011, figs. 1, 6). 19, Falange IV-1 y 20, falange III-2 de Eolambia (McDonald et al., 2012, figs. 35D, K). 21, Falange III-2 y 22, falange IV-3 de Edmontosaurus (Zheng et al., 2011, figs. 1, 6). 23–26, Ungueal pedal de: 23, Gilmoreosaurus (Prieto-Márquez, 2008, fig. J.10A), 24, Edmontosaurus (Zheng et al., 2011, figs. 1, 6), 25, Lapampasaurus (Coria et al., 2012, fig. 9.10) y 26, Brachylophosaurus (Prieto-Márquez, 2008, fig. J.10B). Dibujos sin escala en vista dorsal (1-3, 5, 9, 13, 16-24), medial (8, 12), proximal (6, 10, 14), distal (7, 11, 15-21, 24-26) y lateral (17, 22).
Material suplementario
Tabla A.1. Términos anatómicos del esqueleto axial (según NAA: Baumel y Witmer, 1993).
|
|||||
|
COLUMNA VERTEBRAL/ COLUMNA VERTEBRALIS (NAA) |
colv |
- |
|||
|
Canal vertebral/ Canalis vertebralis (NAA) |
cav |
Neural canal (Lambe, 1920) |
|||
|
Foramen intervertebral / Foramen intervertebrale (NAA) |
fiv |
- |
|||
|
VÉRTEBRA/ VERTEBRAE (NAA) |
vt |
- |
|||
|
Cuerpo vertebral/ Corpus vertebrae (NAA) |
cvt |
Centrum (Lambe, 1920) |
|||
|
Cara articular caudal/ Facie articularis caudalis (NAA) |
fa.c |
- |
|||
|
Cara articular craneal/ Facie articularis cranealis (NAA) |
fa.cr |
- |
|||
|
Cara articular lateral/ Facie lateralis corporis (NAA) |
fla |
- |
|||
|
Concavidad lateral/ Concavitas lateralis (NAA) |
cocl |
- |
|||
|
Eminencia costolateral/ Eminentia costolateralis (NAA) |
ecl |
Parapophysis (Lambe, 1920) |
|||
|
Parapothysis (Davies, 1983) |
|||||
|
Surface for head of rib (Lambe, 1920) |
|||||
|
Fóvea costal/ Fovea costalis (NAA) |
fc |
- |
|||
|
Proceso costal/ Processus costalis (NAA) |
pcos |
Caudal rib (Langstone, 1960) |
|||
|
Sacral rib (Norman, 1986) |
|||||
|
Cara ventral corporal/ Facie ventralis corporis (NAA) |
fvc |
- |
|||
|
Arco vertebral/ Arcus vertebrae (NAA) |
avt |
Neural arch (Prieto-Márquez, 2008) |
|||
|
Pedúnculo vertebral/ Pediculus vertebrae |
pe |
Lamina lateralis arcus (Baumel y Witmer, 1993) |
|||
|
Escotadura craneal del arco/ Incisura cranialis arcus (NAA) |
icr |
- |
|||
|
Escotadura caudal del arco/ Incisura caudalis arcus (NAA) |
ica |
- |
|||
|
Lámina dorsal del arco/ Lamina dorsalis arcus (NAA) |
lad |
- |
|||
|
Proceso espinoso/ Processus spinosus (NAA) |
esp |
Neural spine (Lull y Wrigth, 1942) |
|||
|
Proceso transverso/ Processus transversus vertebrae (NAA) |
tv |
Diapophysis (Lull y Wrigth, 1942) |
|||
|
Diapothysis (Davies, 1983) |
|||||
|
Transverse process (Campione, 2014) |
|||||
|
Fóvea costal/ Fovea costalis (NAA) |
fc |
Diapophysis (Lull y Wrigth, 1942) |
|||
|
Diapothysis (Davies, 1983) |
|||||
|
Surface for tubercle of rib (Lambe, 1920) |
|||||
|
Lámina postcigotransversal/ Lamina postzygotrasnversalis (modificado de Wilson, 1999) |
lpzt |
Postzygodiapophyseal lamina (Wilson, 1999) |
|||
|
Lámina centrotransversal/ Lamina centrotransversalis posterior (modificado de Wilson, 1999) |
lctp |
Posterior centrodiapophyseal lamina (Wilson, 1999) |
|||
|
Lámina costotransversal/ Lamina costrotranversalis (modificado de Wilson, 1999) |
lct |
Paradiapophyseal lamina (Wilson, 1999) |
|||
|
Fosa infratransversal/ Fossa infratransversalis (modificado de Campione, 2014) |
fitv |
Transverse process fossa (Campione, 2014) |
|||
|
Postzygapophyseal centrodiapophyseal |
|||||
|
fossa (Wilson et al., 2011) |
|||||
|
Cigoapófisis caudal/ Zygapophysis caudalis (NAA) |
zca |
Postzygapophysis (Lambe, 1920) |
|||
|
Postzygoapophyseal process (Prieto-Márquez, 2008) |
|||||
|
Post-zygopothysis (Davies, 1983) |
|||||
|
Lámina intrapostcigapofisial/ Lamina intrapostzygoapophysialis (Wilson, 1999) |
ipz |
- |
|||
|
Cigoapófisis craneal/ Zygapophysis cranealis (NAA) |
zcr |
Prezygapophysis (Lambe, 1920) |
|||
|
Pre-zygopothysis (Davies, 1983) |
|||||
|
Cara articular/ Facie articularis (NAA) |
fa |
- |
|||
|
Foramen vertebral/ Foramen vertebrale (NAA) |
fvt |
Neural canal (Lambe, 1920) |
|||
|
Atlas/ Atlas (NAA) |
at |
- |
|||
|
Cuerpo del atlas/ Corpus Atlantis (NAA) |
cat |
Hypocentrum (Lull y Wrigth, 1942) |
|||
|
Intercentrum (Norman, 1986) |
|||||
|
Fosa contiloidea/ Fossa condyloidea (NAA) |
fco |
Surface for basioccipital (Norman, 1986) |
|||
|
Arco del atlas/ Arcus Atlantis (NAA) |
aat |
- |
|||
|
Axis/ Axis (NAA) |
ax |
Axis (Lull y Wrigth, 1942) |
|||
|
Proceso odontoide/ Dens (NAA; Godefroit et al., 1998) |
den |
Odontoid (Lull y Wrigth, 1942) |
|||
|
Intercentro/ Intercentrum (Cuthbertson y Holmes, 2010) |
itc |
Ventral lip (Main et al., 2014) |
|||
|
Vértebra cervical/ Vertebrae cervicales (NAA) |
vtc |
- |
|||
|
Foramen transversal/ Foramen transversarium (NAA) |
ftv |
Transverse foramen (Campione, 2014) |
|||
|
Torus dorsal/ Torus dorsalis (NAA) |
tdo |
Epipophysis (McDonald et al., 2014) |
|||
|
Surco carotídeo/ Sulcus caroticus (NAA) |
suc |
- |
|||
|
Forámenes pneumáticos/ Foramina pneumatica (NAA) |
fpm |
Pleurocoel (Coria et al., 2012) |
|||
|
Vértebra torácica/ Vertebrae thoracicae (NAA) |
vtt |
Dorsal vertebrae (Lambe, 1920) |
|||
|
Cresta ventral corporal/ Crista ventralis corporis (NAA) |
crv |
- |
|||
|
Lámina prespinal/ Lamina prespinalis (Wilson, 1999) |
lpe |
Prezygapophysis septum (Campione, 2014) |
|||
|
Fosa espinopostcigapofisial/ fossa spinopostzygoapophysialis (Wilson, 1999) |
fspo |
Sulcus (Conti et al., 2020) |
|||
|
Fosa espinoprecigoapofisial/ fossa spinoprezygoapophysialis (Wilson, 1999) |
fspe |
Sulcus (Conti et al., 2020) |
|||
|
Hiposfeno/ Hyposphene (McDonald et al., 2014) |
hps |
- |
|||
|
Sacro/ Sacrum (NAA) |
sa |
Sacrum (Lambe, 1920) |
|||
|
Vértebra sacra/ Vertebrae sacri (NAA) |
vts |
Sacral vertebrae (Lambe, 1920) |
|||
|
Vértebra toracosacral/ Vertebrae thoracosacrale (modificado de Prieto-Márquez, 2001) |
vtts |
Dorsosacral vertebrae (Prieto-Márquez, 2001) |
|||
|
Vértebra sacrocaudal/ Vetebrae sacrocaudales (Prieto-Márquez, 2001) |
vtsc |
- |
|||
|
Cara articular iliaca/ Facies articularis ilii |
fa.il |
- |
|||
|
Lámina sacral/ Lamina sacri |
lsa |
- |
|||
|
Yugo sacral/ Jugum sacri |
jsa |
Acetabular bar (Davies, 1983) |
|||
|
Ilia attachment site (Campione, 2014) |
|||||
|
Yoke-like sacral bar (Prieto-Márquez et al., 2012) |
|||||
|
Sacricostal yoke (Norman, 2002) |
|||||
|
Surco ventral corporal/ Sulcus ventralis corporis (NAA) |
suv |
Ventral sulcus (Prieto-Márquez, 2008) |
|||
|
Medial ventral groove (Campione, 2014) |
|||||
|
Ventral groove (Weishampel y Horner, 1990) |
|||||
|
Cresta ventral sacral/ Crista ventralis sacri (NAA) |
cvs |
Crista ventralis synsacri (Baumel y Witmer, 1993) |
|||
|
Ventral ridge (Weishampel y Horner, 1990) |
|||||
|
Vértebra caudal/ Vertebrae caudales (NAA) |
vtca |
- |
|||
|
Hemapófisis/ Hemapophysis (Norman, 1986) |
hp |
Hemal arch articulation site (Campione, 2014) |
|||
|
Chevron facet (Davies, 1983) |
|||||
|
Cresta lateral/ Crista lateralis (Dalla Vecchia, 2020) |
crl |
- |
|||
|
COSTILLA/ COSTAE (NAA) |
cos |
- |
|||
|
Extremo dorsal costal/ Extremitas dorsalis costae (NAA) |
exd |
- |
|||
|
Ángulo costal/ Angulus costae (NAA) |
acs |
- |
|||
|
Capítulo costal/ Capitulum costae (NAA) |
cacs |
Capitulum (Lull y Wrigth, 1942) |
|||
|
Head of the rib (Lambe, 1920) |
|||||
|
Cuello costal/ Collum costae (NAA) |
cucs |
- |
|||
|
Escotadura capitulotubercular/ Incisura capitulotubercularis (NAA) |
itv |
- |
|||
|
Tubérculo costal/ Tuberculum costae (NAA) |
tcs |
Tuberculum (Lull y Wrigth, 1942) |
|||
|
Tubercle (Lambe, 1920) |
|||||
|
Cuerpo costal/ Corpus costae (NAA) |
ccos |
- |
|||
|
Cresta costal/ Crista costae |
crcs |
Ridge (Norman, 1986) |
|||
|
Extremo ventral costal/ Extremitas ventralis costae (NAA) |
exv |
- |
|||
|
Costilla cervical/ Costa cervicale (Lambe, 1920) |
cosc |
- |
|||
|
Costilla torácica/ Costa thoraci (Lambe, 1920) |
cost |
Dorsal rib (Weishampel y Horner, 1990) |
|||
|
CHEURÓN/ CHEVRON (Lambe, 1920) |
ch |
Hemal arch (Prieto-Márquez et al., 2012) |
|||
|
Hemal spine (Campione, 2014) |
|||||
|
Cara articular vertebral/ Facies articularis vertebrae |
fa.v |
Articular facet (McDonald et al., 2012) |
|||
|
Proximal head (Norman, 1986) |
|||||
|
Foramen hemal/ Foramen haemal |
fh |
Haemal canal (Norman, 1986) |
|||
|
Arco hemal/ Arc haemal (Norman, 1986) |
ah |
- |
|||
|
Espina hemal/ Spina haemal (Norman, 1986) |
sh |
- |
|||
|
TENDÓN OSIFICADO/ TENDO OSSIFICANS (Norman, 1986) |
tos |
- |
|||
Tabla A.1 (continuación). Términos anatómicos del esqueleto axial (según NAA: Baumel y Witmer, 1993).
Tabla A.1 (continuación). Términos anatómicos del esqueleto axial (según NAA: Baumel y Witmer, 1993).
Tabla A.2. Términos anatómicos de la cintura pectoral (según NAA: Baumel y Witmer, 1993).
|
Término en español/ latín (referencia) |
Abreviaturas |
Sinónimos en inglés (Referencia) |
|
CINTURA PECTORAL/ CINGULI MEMBRI THORACICI (NAA) |
cmt |
- |
|
Cavidad glenoidea/ Cavitas glenoidalis (NAA) |
cgl |
Glenoid fossa (Davies, 1983) |
|
CORACOIDE/ CORACOIDEUM (NAA) |
cor |
- |
|
Cuerpo coracoideo/ Corpus coracoidei (NAA) |
cco |
- |
|
Cara articular humeral/ Facies articularis humeralis (NAA) |
fa.h |
Glenoid (Brett-Surman, 1989) |
|
Cara articular escapular/ Facies articularis scapularis (NAA) |
fa.sc |
Scapular suture (Brett-Surman yWagner, 2007) |
|
Scapular facet (Davies, 1983) |
||
|
Scapula articulation (Campione, 2014) |
||
|
Tubérculo del bíceps/ Tuberculum biceps (Brett-Surman y Wagner, 2007) |
tubi |
Bicipital knob (Prieto-Márquez y Salinas, 2010) |
|
Lateral process (Brett-Surman y Wagner, 2007) |
||
|
Foramen del nervio supracoracoideo/ Foramen nervous supracoracoidei (NAA) |
fsu |
Coricoid foramen (Davies, 1983) |
|
Coracoid foramen (Brett-Surman, 1989) |
||
|
Extremo esternal coracoideo/ Extremitas sternalis coracoidei (NAA) |
exs |
Hook (Davies, 1983) |
|
Ventral process (Prieto-Márquez, 2008) |
||
|
Ventral hook (Brett-Surman y Wagner, 2007) |
||
|
Sternal blade (McDonald et al., 2012) |
||
|
Hook-like process (Brett-Surman, 1989) |
||
|
Coracoidal hook (Brett-Surman, 1989) |
||
|
Ángulo medial/ Angulus medialis (NAA) |
ame |
- |
|
Cresta articular esternal/ Crista articularis sternalis (NAA) |
cas |
Coracoid ridge (Brett-Surman, 1989) |
|
Coracoidal ridge (Brett-Surman y Wagner, 2007) |
||
|
Escotadura ventral esternal/ Incisura ventralis sternalis |
ivs |
- |
|
ESCÁPULA/ SCAPULA (NAA) |
sc |
- |
|
Extremo craneal escapular/ Extremitas cranialis scapulae (NAA) |
ecr |
Proximal part (Brett-Surman, 1989) |
|
Acromion/ Acromion (NAA) |
acr |
Acromial ridge (Davies, 1983) |
|
Acromion process (Weishampel y Horner, 1990) |
||
|
Pseudoacromion process (Prieto-Márquez y Salinas, 2010) |
||
|
Proceso glenoideo escapular/ Processus glenoidalis scapulae (NAA) |
pgl |
Ventral protuberance (Godefroit et al., 1998) |
|
Scapular labrum (McDonald et al., 2012) |
||
|
Glenoid apex (Prieto-Márquez et al., 2019) |
||
|
Cara articular humeral/ Facies articularis humeralis (NAA) |
fa.h |
Glenoid (Brett-Surman, 1989) |
|
Cara articular coracoidea/ Facies articularis coracoidea (NAA) |
fa.co |
Coracoidal facet (Davies, 1983) |
|
Coracoid articulation (Campione, 2014) |
||
|
Suture for the coracoid (Brett-Surman, 1989) |
||
|
Cresta deltoidea/ Crista deltoidei (Brett-Surman, 1989) |
cdl |
- |
|
Fosa deltoidea/ Fossa deltoidei (Davies, 1983) |
fdl |
- |
|
Cuello escapular/ Collum scapulae (NAA) |
cusc |
Neck (Brett-Surman, 1989) |
|
Proximal constriction (Prieto-Márquez et al., 2019) |
||
|
Cuerpo escapular/ Corpus scapulae (NAA) |
csc |
Blade (Davies, 1983) |
|
Scapular blade (Brett-Surman, 1989) |
||
|
Borde dorsal/ Margo dorsalis (NAA) |
md |
- |
|
Borde ventral/ Margo ventralis (NAA) |
mv |
- |
|
Extremo caudal escapular/ Extremitas caudalis scapulae (NAA) |
eca |
- |
|
PLACA ESTERNAL/ LAMINA STERNALIS (Horner et al., 2004) |
lst |
- |
|
Cuerpo esternal/ Corpus sterni (NAA) |
cst |
Craniomedial plate (Prieto-Márquez, 2008) |
|
Medial flange (Campione, 2014) |
||
|
Sternal blade (McDonald et al., 2012) |
||
|
Sternal paddle (Brett-Surman, 1989) |
||
|
Sternal plate (Brett-Surman y Wagner, 2007) |
||
|
Proximal plate (Godefroit et al., 2012) |
||
|
Proceso caudomedial esternal/ Processus caudomedialis sterni (McDonald et al., 2014) |
pcm |
Caudoventral process (Prieto-Márquez, 2008) |
|
Proceso caudolateral esternal/ Processus caudalteralis sterni (Prieto-Márquez, 2008) |
pcl |
Sternal handle (Brett-Surman, 1989) |
|
Xiphisternum (Brett-Surman y Wagner, 2007) |
Tabla A.2 (continuación). Términos anatómicos de la cintura pectoral (según NAA: Baumel y Witmer, 1993).
Tabla A.3. Términos anatómicos del brazo y antebrazo (según NAA: Baumel y Witmer, 1993).
|
Término en español/ latín (referencia) |
Abreviaturas |
Sinónimos en inglés (Referencia) |
|
HÚMERO/ HUMERUS (NAA) |
h |
- |
|
Extremidad proximal humeral/ Extremitas proximalis humeri (NAA) |
expo |
Dorsal part (Brett-Surman, 1989) |
|
Cabeza humeral/ Caput humeri (NAA) |
ch |
Articular head (Godefroit et al., 1998) |
|
Head (Lambe, 1920) |
||
|
Escotadura de la cabeza humeral/ Incisura capitis humeri (NAA) |
ich |
- |
|
Tubérculo lateral/ Tuberculum lateralis (Davies, 1983) |
tul |
Outer tuberosity (Lambe, 1920) |
|
Tuberculum dorsale (Baumel y Witmer, 1993) |
||
|
Cresta deltopectoral/ Crista deltopectoralis (NAA) |
cdp |
Radial or deltoid crest (Lambe, 1920) |
|
Ángulo de la cresta/ Angulus cristae (NAA) |
agc |
- |
|
Tubérculo medial/ Tuberculum medialis (Davies, 1983) |
tum |
Inner tuberosity (Lambe, 1920) |
|
Tuberculum ventrale (Baumel y Witmer, 1993) |
||
|
Cresta del bíceps/ Crista bicipital (NAA) |
cbi |
- |
|
Plano intertubercular/ Planum intertuberculare (NAA) |
pin |
- |
|
Cuerpo humeral/ Corpus humeri (NAA) |
coh |
Shaft (Brett-Surman, 1989) |
|
Humeral shaft (Brett-Surman y Wagner, 2007) |
||
|
Línea del músculo dorsal ancho/ Linea muscle latissimi dorsi (NAA) |
lida |
Attachment site for latissimus dorsi (Campione, 2014) |
|
Cuello humeral/ Collum humeri |
cuh |
- |
|
Extremo distal humeral/ Extremitas distalis humeri (NAA) |
exdi |
Shaft (Brett-Surman, 1989) |
|
Cóndilo lateral humeral/ Condylus lateralis humeri (modificado de Brett-Surman y Wagner, 2007) |
col |
Lateral condyle (Brett-Surman y Wagner, 2007) |
|
Radial condyle (Norman, 1986) |
||
|
Outer condyle (Lambe, 1920) |
||
|
Condylus dorsalis humeri (Baumel y Witmer, 1993) |
||
|
Cóndilo medial humeral/ Condylus medialis humeri (modificado de Brett-Surman y Wagner, 2007) |
com |
Medial condyle (Brett-Surman y Wagner, 2007) |
|
Ulnar condyle (Norman, 1986) |
||
|
Inner condyle (Lambe, 1920) |
||
|
Condylus ventralis humeri (Baumel y Witmer, 1993) |
||
|
Escotadura intercondilar/ Incisura intercondylaris (NAA) |
iico |
- |
|
Fosa del músculo braquial/ Fossa muscle brachialis (NAA) |
fbr |
- |
|
Epicóndilo lateral/ Epicondylus lateralis (modificado de Baumel y Witmer, 1993) |
epl |
Epicondylus dorsalis (Baumel y Witmer, 1993) |
|
Entepicondylar groove (Campione, 2014) |
||
|
Fosa del olécranon/ Fossa olecrani (NAA) |
fol |
Caudal groove (Prieto-Márquez et al., 2019) |
|
ULNA/ ULNA (NAA) |
ul |
- |
|
Extremo proximal ulnar/ Extremitas proximalis ulnae (NAA) |
expo |
- |
|
Olécranon/ Olecranon (NAA) |
ol |
Olecranon process (Lambe, 1920) |
|
Escotadura del olécranon/ Incisura olecrani (Brett-Surman, 1989) |
iol |
Sigmoidal gutter (Godefroit et al., 1998) |
|
Proximal radial facet (Godefroit et al., 2012a) |
||
|
Cresta lateral/ Crista lateralis (Godefroit et al., 1998) |
crl |
Lateral flange (McDonald et al., 2012) |
|
Cresta medial/ Crista medialis |
crm |
Cranial process (Campione, 2014) |
|
Median process (Godefroit et al., 1998) |
||
|
Medial flange (McDonald et al., 2012) |
||
|
Impresión braquial/ Impressio brachialis (NAA) |
imb |
- |
|
Escotadura radial/ Incisura radialis (NAA) |
Ir |
Radial Surface (Godefroit et al., 1998) |
|
Radial notch (Brett-Surman y Wagner, 2007) |
||
|
Cuerpo ulnar/ Corpus ulnare (NAA) |
coul |
- |
|
Borde caudal/ Margo caudalis (NAA) |
mca |
- |
|
Borde interóseo/ Margo interosseus (NAA) |
mio |
- |
|
Extremo ditsl ulnar/ Extremitas distalis ulnae (NAA) |
exdi |
- |
|
Depresión radial/ Depressio radialis (NAA) |
der |
Distal radial facet (Godefroit et al., 2012a) |
|
Cara articular carpal/ Facies articularis carpalis |
fa.c |
- |
|
RADIO/ RADIUS (NAA) |
r |
- |
|
Extremo proximal radial/ Extremitas proximalis radii (NAA) |
expo |
- |
|
Cabeza radial/ Caput radii (NAA) |
cr |
- |
|
Cótila humeral/ Cotyla humeralis (NAA) |
clh |
Proximal facet (Prieto-Márquez et al., 2019) |
|
Cara articular ulnar/ Facies articularis ulnaris (NAA) |
fa.ul |
- |
|
Cuerpo radial/ Corpus radii (NAA) |
cor |
- |
|
Línea intermuscular/ Lineae intermusculares (NAA) |
lim |
Proximal keel (Godefroit et al., 2012a) |
|
Caudal keel (Godefroit et al., 2012b) |
||
|
Ventral ridge (Prieto-Márquez et al., 2019) |
||
|
Interosseous ridge (Tsogtbataar et al., 2019) |
||
|
Extremo distal radial/ Extremitas distalis radii (NAA) |
exdi |
- |
|
Cara articular radiocarpal/ Facies articularis radiocarpalis (NAA) |
fa.rc |
- |
|
Cara articular ulnar/ Facies articularis ulnaris (NAA) |
fa.ul |
Distal ulnar facet (Godefroit et al., 2012a) |
|
CARPO/ CARPI (NAA) |
cp |
- |
|
Cara articular metacarpal/ Facies articularis metacarpalis (NAA) |
fa.mc |
- |
|
Cara articular radial/ Facies articularis radialis (NAA) |
fa.r |
- |
|
Cara articular ulnar/ Facies articularis ulnaris (NAA) |
fa.ul |
- |
|
Carpiano proximal/ Carpi proximalia (NAA) |
cpp |
- |
|
Carpiano quinto/ Carpi quintus |
cp-5 |
- |
|
Carpiano central/ Carpi centralia (NAA) |
cen |
Intermedium carpal (Tsogtbaatar et al., 2019) |
|
Carpiano radiocentral/ Carpi radialis-centralia |
ra + ce |
- |
|
Carpiano radial/ Carpi radiale (NAA) |
rae |
- |
|
Carpiano ulnar/ Carpi ulnare (NAA) |
ule |
- |
|
Carpiano distal (2-5) / Carpi distalis (2-5) |
cd (2-5) |
- |
|
METACARPO/ METACARPUS (NAA) |
mc |
- |
|
Extremo proximal metacarpal/ Extremitas proximalis metacarpi (modificado de Baumel y Witmer, 1993) |
exp |
- |
|
Cara articular carpal/ Facies articularis carpi |
fa.c |
- |
|
Cuerpo metacarpal/ Corpus metacarpi |
comc |
Metacarpal body (NUEVO) |
|
Cara articular metacarpal/ Facies articularis metacarpi |
fa.m (II-IV) |
Surface for metacarpal II-IV (McDonald et al., 2012) |
|
Cresta medial/ Crista medialis (McDonald et al., 2012) |
cme |
- |
|
Cresta lateral/ Crista lateralis |
cla |
Lateral buttress (McDonald et al., 2012) |
|
Extremo distal metacarpal/ Extremitas distalis metacarpi |
exd |
- |
|
Cara articular digital/ Facies articularis digitorum |
fa.di |
- |
|
Tróclea digital/ Trochlea digitorum |
trdi |
Caudodistal furrow (McDonald et al., 2012) |
|
FALANGE MANUAL/ PHALANX MANUS (Brett-Surman, 1989) |
Ph |
- |
|
Base de la falange/ Basis phalangis |
bph |
- |
|
Cara articular metacarpal/ Facies articularis metacarpalis (NAA) |
fa.m |
- |
|
Cara articular falangeal/ Facies articularis phalangealis (NAA) |
fa.ph |
- |
|
Cuerpo falangeal/ Corpus phalangis (NAA) |
coph |
Shaft (Moreno et al., 2007) |
|
Cabeza falangeal/ Caput phalangis (Moreno et al., 2007) |
cph |
- |
|
Cara articular falangeal/ Facies articularis phalangealis (NAA) |
fa.ph |
- |
|
Tróclea falangeal/ Trochlea phalangealis |
trph |
- |
|
Falange distal/ Phalanx distalis |
phd |
- |
|
Falange media/ Phalanx medius |
phm |
- |
|
Falange proximal/ Phalanx proximalis |
php |
- |
|
Falange ungueal/ Phalanx ungualis (NAA) |
un |
- |
|
Lámina lateroventral/ Lamina lateroventralis |
lalv |
- |
|
Proceso laminar/ Processus laminaris |
pla |
- |
|
Surco neurovascular/ Sulcus neurovascularis (NAA) |
snv |
- |
Tabla A.3 (Continuación). Términos anatómicos del brazo y antebrazo (según NAA: Baumel y Witmer, 1993).
Tabla A.3 (Continuación). Términos anatómicos del brazo y antebrazo (según NAA: Baumel y Witmer, 1993).
Tabla A.4.Términos anatómicos de la cintura pélvica (según NAA: Baumel y Witmer, 1993).
|
Término en español/ latín (referencia) |
Abreviaturas |
Sinónimos en inglés (Referencia) |
|
CINTURA PÉLVICA/ CINGULI MEMBRI PELVICI (NAA) |
cipe |
- |
|
Acetábulo/ Acetabulum (NAA) |
ac |
- |
|
Foramen acetabular/ Foramen acetabuli (NAA) |
fac |
- |
|
Foramen obturador/ Foramen obturatum (NAA) |
fob |
- |
|
Fenestra isquiopúbica/ Fenestra ischiopubica (NAA) |
fisp |
Obturator foramen (Davies, 1983) |
|
Obturator fenestra (Brett-Surman, 1989) |
||
|
Escotadura ilioisquiática/ Incisura Ilioischiadicum (NAA) |
iis |
- |
|
Escotadura iliopúbica/ Incisura ilipubica (NAA) |
iip |
- |
|
ILION/ ILIUM (NAA) |
Il |
- |
|
Ala preacetabular iliaca/ Ala preacetabularis ilii (NAA) |
ape |
Anterior process (Davies, 1983) |
|
Preacetabular process (Brett-Surman, 1989) |
||
|
Cranial process (Weishampel y Horner, 1990) |
||
|
Cresta dorsal iliaca/ Crista iliaca dorsalis (NAA) |
cid |
- |
|
Cresta ventral iliaca/ Crista iliaca ventralis |
civ |
Crista iliaca lateralis (Baumel y Witmer, 1993) |
|
Cresta preacetabular/ Crista preacetabularis |
cpe |
Horizontal boot (McDonald et al., 2017) |
|
Área articular vertebral/ Area articulares vertebrales (NAA) |
aav |
Medial sacral ridge (Prieto-Márquez, 2008) |
|
Medial shelf (Carpenter e Ishida, 2010) |
||
|
Escotadura preacetabular/ Incisura preacetabularis (Godefroit et al., 1998) |
ipe |
- |
|
Ala postacetabular iliaca/ Ala postacetabularis ilii (NAA) |
apo |
Posterior process (Davies, 1983) |
|
Postacetabular process (Brett-Surman, 1989) |
||
|
Caudal process (Weishampel y Horner, 1990) |
||
|
Cara dorsal/ Facies dorsalis (NAA) |
fd |
- |
|
Cresta caudodorsal/ Crista caudodorsalis |
ccd |
Median ridge (Prieto-Márquez, 2008) |
|
Dorsal ridge (Prieto-Márquez y Salinas, 2010) |
||
|
Cresta dorsolateral iliaca/ Crista dorsolaterallis illi (NAA) |
cdil |
Lateral iliac crest (Carpenter e Ishida, 2010) |
|
Suprailiac crest (Carpenter e Ishida, 2010) |
||
|
Ridge of supracetabular process (Prieto-Márquez, 2008) |
||
|
Cara lateral/ Facies lateralis (NAA) |
fla |
- |
|
Concavidad infracristal/ Concavitas infracristalis (NAA) |
cnif |
- |
|
Fosa breve/ Fossa brevis (Norman, 1986) |
fb |
Brevis fossa (Norman, 1986) |
|
Brevis shelf (Norman, 1986) |
||
|
Cara medial/ Facies medialis (NAA) |
fme |
- |
|
Cresta breve/ Crista brevis (modificado de Prieto-Márquez, 2008) |
cbm |
Medial ridge (Prieto-Márquez, 2008) |
|
Escotadura postacetabular/ Incisura postacetabularis (Godefroit et al., 1998) |
ipo |
- |
|
Cuerpo iliaco/ Corpus ilii (NAA) |
coil |
Central plate (Prieto-Márquez, 2008) |
|
Main plate (Godefroit et al., 1998) |
||
|
Mid-section (Brett-Surman, 1989) |
||
|
Iliac plate (Norman, 1986) |
||
|
Body (Carpenter e Ishida, 2010) |
||
|
Proceso lateral iliaco/ Processus lateralis illi (NAA y Carpenter e Ishida, 2010) |
pli |
Antitrochanter (Lull y Wrigth, 1942) |
|
Iliac boss (Norman, 2014) |
||
|
Supraacetabular process (Prieto-Márquez, 2008) |
||
|
Dorsal flange (McDonald et al., 2017) |
||
|
Suprailiac crest (Brett-Surman, 1989) |
||
|
Laterally everted rim (McDonald et al., 2014) |
||
|
Supratrochanteric crest (Maidment y Barrett, 2012) |
||
|
Escotadura suprailiaca/ Incisura suprailii (Carpenter e Ishida, 2010) |
isil |
- |
|
Fosa iliaca dorsal/ Fossa iliaca dorsalis (NAA) |
fid |
- |
|
Pedúnculo púbico/ Pediculus pubis (Davies, 1983) |
ppu |
Pubic process (Gates et al., 2018) |
|
Cresta supraacetabular/ Crista supraacetabularis |
csaa |
Supra-acetabular flange (Maidment y Barrett, 2011) |
|
Escotadura acetabular/ Incisura acetabularis (NAA) |
iac |
Acetabular surface (Davies, 1983) |
|
Pedúnculo isquiático/ Pediculus ischii (Davies, 1983) |
pis |
Ischial process (Gates et al., 2018) |
|
Ischiadic process (Prieto-Márquez et al., 2019) |
||
|
Antitrocánter/ Antitrochanter (NAA) |
ant |
Caudal tuberosity (Prieto-Márquez, 2008) |
|
Postacetabular ridge (Campione, 2014) |
||
|
Ischial tuberosity (Brett-Surman y Wagner, 2007) |
||
|
Posteroventral protuberance (Gates et al., 2018) |
||
|
Ischiadic tuberosity (Prieto-Márquez et al., 2019) |
||
|
PUBIS/ PUBIS (NAA) |
pu |
- |
|
Cuerpo púbico/ Corpus pubis (NAA) |
copu |
- |
|
Escotadura acetabular/ Incisura acetabularis (NAA) |
iac |
Acetabular margin (Brett-Surman, 1989) |
|
Cresta acetabular/ Crista acetabularis |
cac |
Acetabular margin (Prieto-Márquez, 2008) |
|
Pedúnculo iliaco/ Pediculus illii (Davies, 1983) |
pil |
Dorsal process (Godefroit et al., 1998) |
|
Iliac process (Gates et al., 2018) |
||
|
Pedúnculo isquiático/ Pediculus ischii (Davies, 1983) |
pis |
Ischial process (Gates et al., 2018) |
|
Tuberosidad lateroventral/ Tuberositas lateroventralis |
tulv |
Lateroventral protuberance (Prieto-Márquez, 2008) |
|
Ala prepúbica/ Ala prepubis |
pepu |
Prepubis (Davies, 1983) |
|
Anterior pubic process (Norman, 1986) |
||
|
Prepubic process (Horner et al., 2004) |
||
|
Prepubic part (Brett-Surman, 1989) |
||
|
Cranial pubic process (McDonald et al., 2012) |
||
|
Cuello púbico/ Collum pubis (Brett-Surman, 1989) |
cupu |
Prepubic constriction (Prieto-Márquez, 2008) |
|
Neck (Davies, 1983) |
||
|
Lámina púbica/ Lamina pubis |
lpu |
Pubic blade (Brett-Surman, 1989) |
|
Blade (Davies, 1983) |
||
|
Expansión dorsal/ Expansion dorsalis (Godefroit et al., 2008) |
ed |
- |
|
Expansión ventral/ Expansion ventralis (Godefroit et al., 2008) |
ev |
- |
|
Escapo púbico/ Scapus pubis (NAA) |
spu |
Postpubis (Davies, 1983) |
|
Postpubic (Lull y Wrigth, 1942) |
||
|
Postpubic rod (Norman, 1986) |
||
|
Postpubic process (Brett-Surman, 1989) |
||
|
Pubic bar (Godefroit et al., 1998) |
||
|
Caudal pubic process (McDonald et al., 2012) |
||
|
Foramen obturador/ Obturator foramen (Norman, 1869) |
fob |
- |
|
Escotadura obturadora/ Incisura obturatorius |
iob |
Obturator gutter (Godefroit et al., 1998) |
|
Proceso caudodorsal/ Processus caudodorsalis (McDonald et al., 2012) |
prcd |
Triangular process (Norman, 1869) |
|
Cresta dorsomedial/ Crista dorsomedialis |
cdm |
Dorsal shelf (Ramírez-Velasco et al., 2012) |
|
Surco obturador/ Sulcus obturatorii (NAA) |
sob |
- |
|
ISQUION/ ISCHIUM (NAA) |
is |
- |
|
Cuerpo isquiático/ Corpus ischii (NAA) |
cois |
- |
|
Escotadura acetabular/ Incisura acetabularis (NAA) |
iac |
Acetabulum (Brett-Surman, 1989) |
|
Pedúnculo iliaco/ Pediculus iii (Davies, 1983) |
pil |
Iliac process (Horner et al., 2004) |
|
Pedúnculo púbico/ Pediculus pubis (Davies, 1983) |
ppu |
Pubic process (Horner et al., 2004) |
|
Escotadura isquipúbica/ Incisura ischiopubica |
iisp |
Obturator foramen (Davies, 1983) |
|
Obturator fenetra (Brett-Surman, 1989) |
||
|
Foramen isquiático/ Foramen ischii |
fis |
Obturador gutter (Godefroit et al., 1998) |
|
Proceso obturador/ Processus obturatorius (NAA) |
prob |
Ventral process (Campione, 2014) |
|
Proceso ventral/ Processus ventralis (Horner et al., 2004) |
prv |
Obturador process (Brett-Surman, 1989) |
|
Hook spur (Brett-Surman y Wagner, 2007) |
||
|
Escapo isquiático/ Scapus ischii |
sis |
Ischial shaft (Brett-Surman, 1989) |
|
Shaft (Davies, 1983) |
||
|
Ala ischii (Baumel y Witmer, 1993) |
||
|
Ápice isquiático/ Apex ischii |
ais |
Apex pubis (Baumel y Witmer, 1993) |
|
Bota isquiática/ Pes ischii (Brett-Surman, 1989) |
pei |
Foot (Brett-Surman y Wagner, 2007) |
|
Sínfisis isquiática/ Symphysis ischii |
sin |
- |
|
Depresión lateral/ Depressio lateralis (Conti et al., 2019) |
dla |
- |
Tabla A.4 (Continuación).Términos anatómicos de la cintura pélvica (según NAA: Baumel y Witmer, 1993).
Tabla A.4 (Continuación).Términos anatómicos de la cintura pélvica (según NAA: Baumel y Witmer, 1993).
Tabla A.5. Términos anatómicos del miembro pélvico (según NAA: Baumel y Witmer, 1993).
|
Término en español/ latín (referencia) |
Abreviaturas |
Sinónimos en inglés (Referencia) |
|
FÉMUR/ FEMUR (NAA) |
fe |
- |
|
Extremo proximal femoral/ Extremitas proximalis femoris (NAA) |
expo |
- |
|
Cabeza femoral/ Caput femoris (NAA) |
cfe |
Head (Davies, 1983) |
|
Cuello femoral/ Collum femoris (NAA) |
cufe |
Neck of femoral head (McDonald et al., 2012) |
|
Trocánter craneal/ Trochanter cranialis (Horner et al., 2004) |
trc |
Lesser trochanter (Davies, 1983) |
|
Trochanter femoris (Baumel y Witmer, 1993) |
||
|
Trocánter mayor/ Trochanter mayoris (Davies, 1983) |
trm |
Trochanter femoris (Baumel y Witmer, 1993) |
|
Cuerpo femoral/ Corpus femoris (NAA) |
cofe |
- |
|
Cuarto trocánter/ Trochanter quarto (Lull y Wrigth, 1942) |
4tr |
- |
|
Impresión del músculo caudofemoral/ Impression muscle caudofemoralis |
icf |
Attachment site for caudofemoralis longus (Campione, 2014) |
|
Impresión del músculo puboisquiofemoral/ Impression muscle puboischiofemoralis (NAA) |
ipif |
Attachment site for puboischiofemoralis internus (Campione, 2014) |
|
Impresión del músculo femorotibial/ Impression muscle femorotibialis (NAA) |
ift |
- |
|
Extremo distal femoral/ Extremitas distalis femoris (NAA) |
exdi |
- |
|
Cóndilo lateral/ Condylus lateralis (NAA) |
col |
- |
|
Tróclea fibular/ Trochlea fibularis (NAA) |
ctifi |
- |
|
Cóndilo medial/ Condylus medialis (NAA) |
com |
- |
|
Surco intercondilar/ Sulcus intercondylaris (NAA) |
suic |
Intercondylar groove (Davies, 1983) |
|
Extensor tunnel (Campione, 2014) |
||
|
Cranial depression (Campione, 2014) |
||
|
Anterior intercondylar groove (Godefroit et al., 1998) |
||
|
Condylar canal (Brett-Surman, 1989) |
||
|
Fosa poplítea/ Fossa poplítea (NAA) |
fpo |
Posterior intercondylar groove (Godefroit et al., 1998) |
|
Caudal depression (Campione, 2014) |
||
|
TIBIA/ TIBIA (NAA) |
ti |
- |
|
Extremo proximal tibial/ Extremitas proximalis tibiae (modificado de Baumel y Witmer, 1993) |
expo |
Extremitas proximalis tibiotarsi (Baumel y Witmer, 1993) |
|
Cresta cnemial/ Crista cnemialis (Davies, 1983) |
ccm |
Crista cnemialis lateralis (Baumel y Witmer, 1993) |
|
Cóndilo lateral/ condylus lateralis (McDonald et al., 2012) |
col |
Inner condyle (Norman, 1986) |
|
Cóndilo medial/ condylus medialis |
com |
Outer condyle (Norman, 1986) |
|
Caudomedial condyle (McDonald et al., 2012) |
||
|
Área interarticular/ Area interarticularis (NAA) |
air |
Intercondylar groove (Norman, 1986) |
|
Cuerpo tibial/ Corpus tibial (modificado de Baumel y Witmer, 1993) |
coti |
Corpus tibiotarsi (Baumel y Witmer, 1993) |
|
Borde lateral/ Margo lateralis (NAA) |
mla |
- |
|
Borde medial/ Margo medialis (NAA) |
mme |
- |
|
Extremo distal tibial/ Extremitas distalis tibialis (Brett-Surman, 1989) |
exdi |
Extremitas distalis tibiotarsi (Baumel y Witmer, 1993) |
|
Surco extensor/ Sulcus externsorius (NAA) |
sex |
- |
|
Maleólo lateral/ Malleoulus lateralis (Campione, 2014) |
mll |
External malleolus (Godefroit et al., 1998) |
|
Outer malleolus (Davies, 1983) |
||
|
Hemimalleoulus tibial (Ramírez-Velasco et al., 2021) |
||
|
Cara articular fibular/ Facies articularis fibularis |
fa.fi |
Fibular contact (Norman, 1986) |
|
Concavity for articulation of distal end of fibula (McDonald et al., 2012) |
||
|
Maleólo medial tibial/ Malleolus medialis (Campione, 2014) |
mlm |
Internal malleolus (Godefroit et al., 1998) |
|
Inner malleoulus (Davies, 1983) |
||
|
Tróclea del cartílago tibial/ Trochlea cartilaginis tibialis (NAA) |
trct |
- |
|
FÍBULA/ FIBULA (NAA) |
fi |
- |
|
Cabeza fibular/ Caput fibulae (NAA) |
cfi |
- |
|
Cara articular femoral/ Facies articularis femoralis (NAA) |
fa.fe |
- |
|
Cara articular tibial/ Facies articularis tibialis (NAA) |
fa.ti |
Concave surface for articulation with lateral and caudomedial condyles for tibia (McDonald et al., 2012) |
|
Ápice craneal/ Apex cranialis |
axc |
Anterior process (Prieto-Márquez y Norell, 2010) |
|
Cranial process (Prieto-Márquez, 2014) |
||
|
Cranial wing on proximal end of fibula (McDonald et al., 2012) |
||
|
Cuerpo fibular/ Corpus fibulae (NAA) |
cofi |
- |
|
Extremo distal/ Extremitas distalis |
exdi |
- |
|
Proceso distal/ processus distalis (Prieto-Márquez y Norell, 2010) |
pdi |
Distal end (Prieto-Márquez, 2008) |
|
Fibular foot (Norman, 1986) |
||
|
Cranial expansion of distal end of fibula (McDonald et al., 2012) |
||
|
Cara articular tibial/ Facies articularis tibialis (NAA) |
fa.ti |
Articular surface for fibula (Prieto-Márquez y Norell, 2010) |
|
Cara articular tarsal/ Facies articularis tarsali (NAA) |
fa.ta |
- |
|
CALCÁNEAO/ CALCANEUM (Norman, 1986) |
ca |
Fibulare (Baumel y Witmer, 1993) |
|
Cresta tibiofibular/ Crista tibiofibularis |
tifi |
Ascending process (Godefroit et al., 1998) |
|
Posterolateral process (Prieto-Márquez y Norell, 2010) |
||
|
Depresión epicondilar lateral/ Depression epicondylaris lateralis (NAA) |
del |
- |
|
Cara articular fibular/ Facies articularis fibularis |
fa.fi |
Fibular facet (Godefroit et al., 1998) |
|
Cara articular tibial/ Facies articularis tibialis |
fa.ti |
Tibial facet (Godefroit et al., 1998) |
|
Cóndilo lateral/ Condylus lateralis (NAA) |
col |
- |
|
ASTRÁGALO/ ASTRAGALUS (Norman, 1986) |
as |
Tibiale (Baumel y Witmer, 1993) |
|
Cara articular tibial/ Facies articularis tibialis |
fa.ti |
Articular surface for the tibia (Godefroit et al., 1998) |
|
Cóndilo medial/ Condylus medialis (NAA) |
com |
Medial platform (Prieto-Márquez, 2008) |
|
Escotadura intercondilar/ Incisura intercondylaris (NAA) |
iic |
- |
|
Proceso ascendente/ Processus ascendens (Campione, 2014) |
asc |
Anterior ascending process (Brett-Surman, 1989) |
|
Cóndilo lateral/ Condylus lateralis (NAA) |
col |
- |
|
Depresión intercondilar/ Depression intercondylaris |
dea |
- |
|
TARSAL DISTAL/ TASI DISTALE (NAA) |
td |
- |
|
METATARSO/ METATARSALE (NAA) |
mt |
- |
|
Extremo proximal metatarsal/ Extremitas proximalis metatarsi (modificado de Baumel y Witmer, 1993) |
exp |
Extremitas proximalis tarsometatarsi (Baumel y Witmer, 1993) |
|
Proximal articulation process (Prieto-Márquez y Norell, 2010) |
||
|
Cótila articular/ Cotyla articularis (NAA) |
coa |
- |
|
Cresta plantar metatarsal/ Crista plantaris metatarsalis (modificado de Baumel y Witmer, 1993) |
cpm |
Crista plantaris lateralis, medialis (Baumel y Witmer, 1993) |
|
Craneolateral flange (McDonald et al., 2012) |
||
|
Craneomedial flange (McDonald et al., 2012) |
||
|
Medioventral flange (Prieto-Márquez, 2010) |
||
|
Medial flange (Prieto-Márquez et al., 2019) |
||
|
Cresta dorsal metatarsal/ Crista dorsalis metatarsalis |
cdm |
- |
|
Cara articular metatarsal (II-IV) / Facies articularis metatarsalis (II-IV) |
fa.mt II-IV |
- |
|
Cuerpo metatarsal/ Corpus metatarsalis |
comt |
- |
|
Lámina mediodorsal/ Lamina mediodorsalis (modificado de Prieto-Márquez, 2010) |
lam |
Lateral knob (McDonald et al., 2012) |
|
Mediodorsal flange (Prieto-Márquez, 2010) |
||
|
Tuberosidad medial/ Tuberositas medialis (Prieto-Márquez, 2010) |
tum |
Craneomedial knob (McDonald et al., 2012) |
|
Extremo distal metatarsal/ Extremitas distalis metatarsalis (modificado de Baumel y Witmer, 1993) |
exd |
- |
|
Tróclea metatarsal (II-IV) / Trochlea metatarsi (II-IV) (NAA) |
tmt |
Distal articulation process (Prieto-Márquez y Norell, 2010) |
|
Medial condyle and lateral condyle (McDonald et al., 2012) |
||
|
Fóvea del ligamento colateral/ Foveae ligamentorum collateralium (NAA) |
flc |
- |
|
FALANGE PEDAL/ PHALANGES PEDAL (Brett-Surman, 1989) |
ph |
- |
|
Base falangeal/ Basis phalangis (NAA) |
bph |
- |
|
Cótila articular/ Cotyla articularis (NAA) |
coa |
Facies articularis metacarpalis (Baumel y Witmer, 1993) |
|
Proximal joint Surface (Moreno et al., 2007) |
||
|
Cuerpo falangeal/ Corpus phalangis (NAA) |
coph |
Shaft (Moreno et al., 2007) |
|
Cabeza falangeal/ Capitulum phalangis (NAA) |
cph |
Facies articularis phalangealis (Baumel y Witmer, 1993) |
|
Distal joint Surface (Moreno et al., 2007) |
||
|
Tróclea articular/ Trochlea articularis (NAA) |
tar |
- |
|
Fóvea del ligamento colateral/ Foveae ligamentorum collateralium (NAA) |
flc |
- |
|
Falange distal/ Phalanx distalis |
phd |
- |
|
Falange media/ Phalanx medium |
phm |
- |
|
Falange proximal/ Phalanx proximalis |
php |
- |
|
Falange ungueal/ Phalanx ungualis (NAA) |
un |
- |
|
Quilla plantar/ Carinalis plantae |
capl |
Axial keel (Brett-Surman y Wagner, 2007) |
|
Lámina lateroventral/ Lamina lateroventralis |
lalv |
- |
|
Proceso laminar/ Processus laminaris |
pla |
- |
|
Surco neurovascular/ Sulcus neurovascularis (NAA) |
snv |
- |
Tabla A.5 (continuación). Términos anatómicos del miembro pélvico (según NAA: Baumel y Witmer, 1993).
Tabla A.5 (continuación). Términos anatómicos del miembro pélvico (según NAA: Baumel y Witmer, 1993).
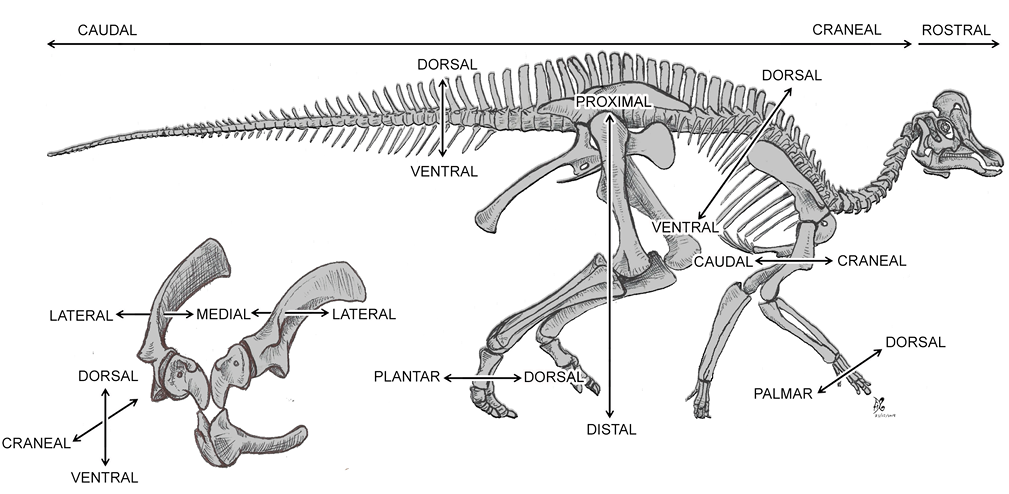
Figura S1. Términos de dirección anatómica empleados en este trabajo. Tomados de la Nomina Anatomica Avium, 1993.
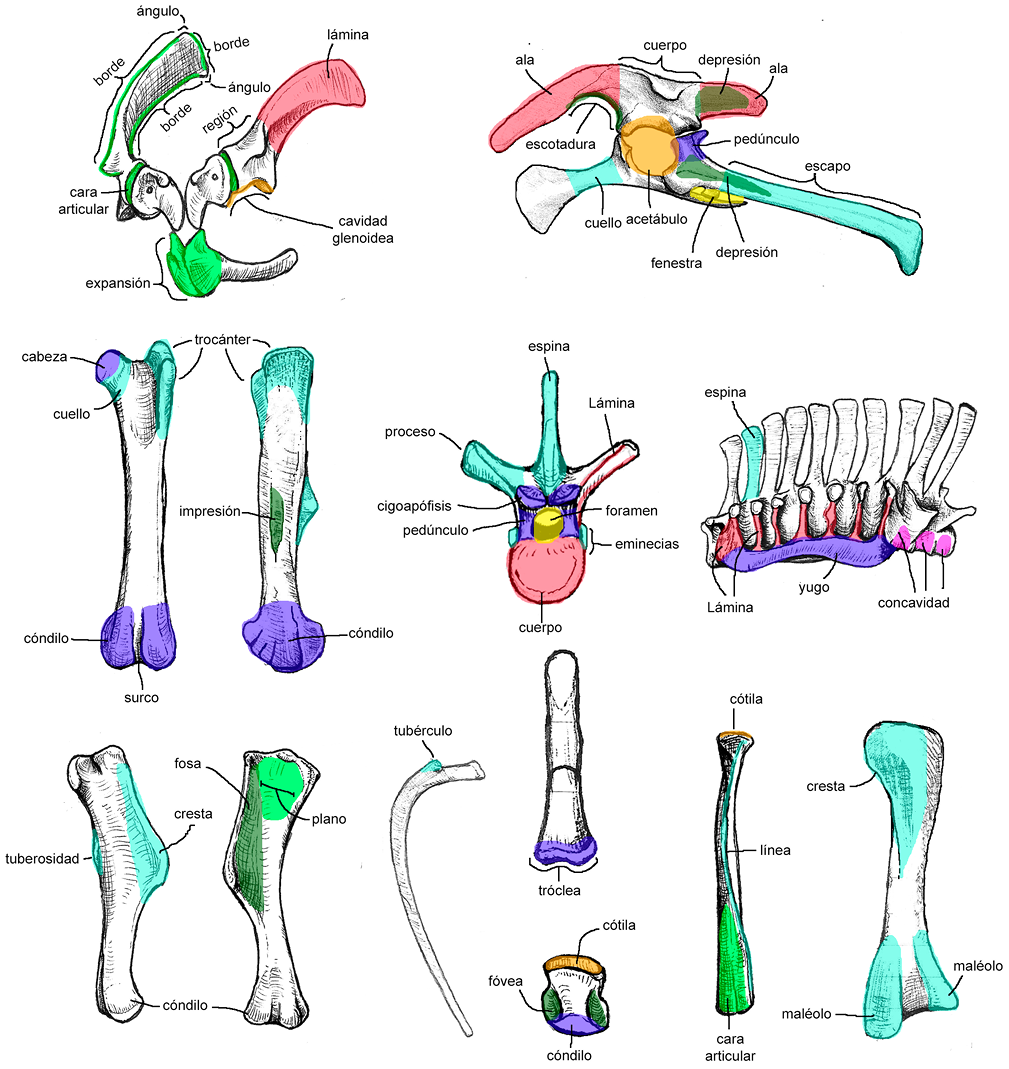
Figura S2. Términos anatómicos utilizados para la descripción de estructuras anatómicas de huesos postcraneanos. El color verde claro indica términos de partes del hueso; color rojo, términos de forma; azul obscuro, proyecciones articulares; azul claro, proyecciones no articulares; naranja, depresiones articulares; verde obscuro, depresiones no articulares; amarillo, huecos. Para definiciones de los términos ver tabla 2.
Referencias suplementarias
Baumel, J.J., Witmer, L.M., 1993, Osteologia, en Baumel, J.J., King, A., Lucas Breazile, A., Evans, H. (eds.), Handbook of avian anatomy: nomina anatomica avium: Cambridge, Massachusetts, Publication of the nuttal ornithological club, 45–136.
Brett-Surman, M.K., 1989, A revision of the Hadrosauridae (Reptilia: Ornithischia) and their evolution during the Campanian and Maastrichtian: Washington, George Washington University, Tesis dictoral, 192 pp.
Brett-Surman, M.K., Wagner, J.R., 2007, Discussion of character analysis of the appendicular anatomy in Campanian and Maastrichtian North America hadrosaurids-variation and ontogeny, en Carpenter, K. (ed.), Horns and beaks: ceratopsian and ornithopod dinosaurs: Bloomington, Indiana University Press, 135–169.
Campione, N., 2014, Postcranial anatomy of Edmontosaurus regalis (Hadrosauridae) from the Horseshoe Canyon Formation, Alberta, Canada, en Eberth, D.A., Evans, D.C. (eds.), Hadrosaurs: Bloomington, Indiana University Press, 208–244.
Carpenter, K., Ishida, Y., 2010, Early and “middle” Cretaceous iguanodonts in time and space: Journal of Iberian Geology, 36, 145–164.
Conti, S., Vila, B., Sellés, A.G., Galobart, A., Benton, M.J., Prieto-Márquez, A., 2020, The oldest lambeosaurine dinosaur from Europe: insights into the arrival of Tsintaosaurini: Cretaceous Research, 107, 104286, doi: 10.1016/j.cretres.2019.104286
Coria, R.A., González Riga, B., Casadio, S., 2012, Un nuevo hadrosáurido (Dinosauria, Ornithopoda) de la Formación Allen, provincia de La Pampa, Argentina: Ameghiniana, 49(4), 552–572, doi: 10.5710/ AMGH.9.4.2012.487
Cuthbertson, R.S., Holmes, R.B., 2010, The first complete description of the holotype of Brachylophosaurus canadensis Sternberg, 1953 (Dinosauria: Hadrosauridae) with comments on intraspecific variation: Zoological Journal of the Linnean Society, 159, 373–397.
Dalla Vecchia, F.M., 2020, The unusual tail of Tethyshadros insularis (Dinosauria, Hadrosauroidea) from the Adriatic Island of the European archipelago: Revista Italiana di Paleontologia e Stratigrafia, 126, 583–628.
Davies, K.L.B.A., 1983, Hadrosaurian dinosaurs of Big Bend National Park Brewster county, Texas: Austin, University of Texas, Tesis de maestría, 235 pp.
Gates, T.A., Tsogtbaatar, K., Zanno, L.E., Chinzoring, T., Watabe, M., 2018, A new iguanodontian (Dinosauria: Ornithopoda) from the Early Cretaceous of Mongolia: PeerJ, 6, e5300, doi: 10.7717/peerj.5300
Godefroit, P., Bolotsky, Y.L., Lauters, P., 2012a, A new saurolophine dinosaur from the latest Cretaceous of Far Eastern Russia: PLoS ONE, 7, e36849, doi: 10.1371/journal.pone.0036849
Godefroit, P., Bolotsky, Y.L., Bolotsky, I.Y., 2012b, Osteology and relationships of Olorotitan arharensis, a hollow−crested hadrosaurid dinosaur from the latest Cretaceous of Far Eastern Russia: Acta Palaeontologica Polonica, 57, 527–560.
Godefroit, P., Hai, S., Yu, T., Lauters, P., 2008, New hadrosaurid dinosaurs from the uppermost Cretaceous of north−eastern China: Acta Palaeontologica Polonica, 53, 47–74, doi, 10.4202/app.2008.0103
Godefroit, P., Dong, Z.M., Bultynck, P., Li, H., Feng, L., 1998, New Bactrosaurus (Dinosauria: Hadrosauroidea) material from Iren Dabasu (Inner Mongolia, P.R. China): Sciences de la Terre, 68, 3–70.
Horner, J.R., Weishampel, D.B., Forster, C.A., 2004. Hadrosauridae, in Weishampel, D.B., Dodson, P., Osmólska, H. (eds.), The dinosauria, second edition: Berkeley, University of California Press, 438–463.
Lambe, L.M., 1920. The hadrosaur Edmontosaurus from the Upper Cretaceous of Alberta, Memoir, 120: Department of Mines, Geological Survey of Canada, 1–79.
Langstone, W.Jr., 1960, The vertebrate fauna of the Selma Formation of Alabama. Part VI. The dinosaurs: Fieldiana: Geology Memoirs, 3, 317–361.
Lull, R.S., Wrigth, N.E., 1942, Hadrosaurian dinosaurs of North America: Geological Society of America, Special Papers no 40, Baltimore, 242 pp.
Main, D.J., Noto, C.R., Weishampel, D.B., 2014, Postcranial anatomy of a basal hadrosauroid (Dinosauria: Ornithopoda) from the Cretaceous (Cenomanian) Woodbine Formation of North Texas, en Eberth, D.A., Evans, D.C. (eds.), Hadrosaurs: Bloomington, Indiana University Press, 77–95.
Maidment, S.C.R., Barrett, P.M., 2011, The locomotor musculature of basal ornithischian dinosaurs: Journal of Vertebrate Paleontology, 31, 1265–1291.
Maidment, S.C.R., Barrett, P.M., 2012, Does morphological convergence imply functional similarity? A test using the evolution of quadrupedalism in ornithischian dinosaurs: Proceeding of the Royal Society B, 279, 3765–3771, doi: 10.1098/rspb.2012.1040
McDonald, A.T., Bird, J., Kirkland, J.I., Dodson, P., 2012, Osteology of the basal hadrosauroid Eolambia caroljonesa (Dinosauria: Ornithopoda) from Cedar Mountain Formation of Utah: PLoS ONE, 7, e45712, doi: 10.1371/journal.pone.0045712
McDonald, A.T., Gates, T.A., Zanno, L.E., Makovicky, P.J., 2017, Anatomy, taphonomy, and phylogenetic implications of a new specimen of Eolambia caroljonesa (Dinosauria: Ornithopoda) from the Cedar Mountain Formation, Utah, USA, PLoS ONE, 12, e0176896, doi: 10.1371/journal.pone.0176896
McDonald, A.T., Maidment, S.C.R., Barrett, P.M., You, H.I., Dodson, P., 2014, Osteology of the basal hadrosauroid Equijubus normani (Dinosauria, Ornithopoda) from the Early Cretaceous of China, in Eberth, D.A., Evans, D.C. (eds.), Hadrosaurs: Bloomington, Indiana University Press, 44–72.
Moreno, K., Carrano, M., Snyder, R., 2007, Morphological changes in pedal phalanges through ornithopod dinosaur evolution: a biomechanical approach: Journal of Morphology, 63, 50–63.
Norman, D.B., 1986, On the anatomy of Iguanodon atherfieldensis (Ornithischia: Ornithopoda): Bulletin de l’Institut royal des Sciences naturelles de Belgique, Sciences de la Terre, 56, 281–372.
Norman, D.B., 2002, On asian ornithopods (Dinosauria: Ornithischia). 4. Probactrosaurus Rozhdestvensky, 1966: Zoological Journal of the Linnean Society, 136, 113–144.
Norman, D.B., 2014, Iguanodonts from the Wealden of England: do they contribute to the discussion concerning hadrosaur origin?, en Eberth, D.A., Evans, D.C. (eds.), Hadrosaurs: Bloomington, Indiana University Press, 10–43.
Prieto-Márquez, A., 2001, Osteology and variation of Brachylophosaurus canadensis (Dinosauria, Hadrosauridae) from the Upper Cretaceous Judith River Formation of Montana: Bozeman, Montana, Montana State University, Tesis de Maestría, 390 pp.
Prieto-Márquez, A., 2008, Phylogeny and historical biogeography of hadrosaurid dinosaurs: Florida, Florida State University College of arts and science, Tesis de doctorado, 861 pp.
Prieto-Márquez, A., 2010, The braincase and skull roof of Gryposaurus notabilis (Dinosauria, Hadrosauridae), with a taxonomic revisión of the genus: Journal of Vertebrate Paleontology, 30, 838–854.
Prieto-Márquez, A., 2014, A juvenile Edmontosaurus from the Late Maastrichtian (Cretaceous) of North America: implications for ontogeny and phylogenetic inference in saurolophine dinosaurs: Cretaceous Research, 50, 282–303, doi: org/10.1016/j. cretres.2014.05.003
Prieto-Márquez, A., Norell, M., 2010, Anatomy and relationships of Gilmoreosaurus mongoliensis (Dinosauria, Hadrosauroidea) from the late Cretaceous of Central Asia: American Museum Novitates, 3694, 1–49, doi: 10.1206/3694.2
Prieto-Márquez, A., Salinas, G.C., 2010, A re-evaluation of Secernosaurus koerneri and Kritosaurus australis (Dinosauria, Hadrosauridae) from the Late Cretaceous of Argentina: Journal of Vertebrate Paleontology, 30, 813–837, doi: 10.1080/02724631003763508
Prieto-Márquez, A., Chiappe, L.M., Joshi, S.H., 2012, The lambeosaurine dinosaur Magnapaulia laticaudus from the Late Cretaceous of Baja California, Northwestern Mexico: PLoS ONE, 7, e38207, doi: 10.1371/journal.pone.0038207
Prieto-Márquez, A., Fondevilla, V., Sellés, A. G., Wagner, J. R., Galobart, À., 2019b, Adynomosaurus arcanus, a new lambeosaurine dinosaur from the Late Cretaceous Ibero-Armorican Island of the European Archipelago: Cretaceous Research, doi: 10.1016/j.cretres.2018.12.002
Ramírez-Velasco, Á.A., Espinosa-Arrubarrena, L., Alvarado-Ortega, J., 2021, Review of the taxonomic affinities of Latirhinus uitstlani, an emblematic Mexican hadrosaurid: Journal of South American Earth Sciences, doi: 10.1016/j.jsames.2021.103391
Ramírez-Velasco, A.A., Benammi, M., Prieto-Márquez, A., Alvarado Ortega, J., Hernández-Rivera, R., 2012, Huehuecanauhtlus tiquichensis, a new hadrosauroid dinosaur (Ornithischia: Ornithopoda) from the Santonian (Late Cretaceous) of Michoacán, Mexico: Canadian Journal of Earth Sciences, 49, 379–395, doi: 10.1139/e11-062
Tsogtbataar, K., Weishampel, D.B., Evans, D.C., Watabe, M., 2019, A new hadrosauroid (Dinosauria: Ornithopoda) from the Late Cretaceous Baynshire Formation of the Gobi Desert (Mongolia): PLoS ONE, 14, e0208480, doi: 10.1371/journal.pone.0208480
Weishampel, D.B., Horner, J.R., 1990, Hadrosauridae, en Weishampel, D.B., Dodson, P., Osmólska, H. (eds), The Dinosauria: Berkeley, University of California Press, 534–551.
Wilson, J.A., 1999, A nomenclatura for vertebral laminae in sauropods and other saurischian dinosaurs: Journal of Vertebrate Paleontology, 19, 639–653.
Wilson, J.A., D’Emic, M.D., Ikejiri, T., Moacdieh, E.M., Whitlock, J.A., 2011, A nomenclature for vertebral fossae in sauropods and other saurischian dinosaurs: PLoS ONE, 6, e17114, doi: 10.1371/journal.pone.0017114